
1.3 生物衰老学家如何研究衰老:实验室生物在人类衰老研究中的应用
伦理和实践方面的考虑限制了可以在人类身上进行的研究类型。因此,生物衰老学家利用多种生物,包括单细胞生物、昆虫在内的无脊椎动物、一系列哺乳动物和鱼类、鸟类、非人灵长类以及一些人类遗传疾病,来研究人类衰老的基本性质。本节简要介绍了作为研究人类衰老和长寿机制的实验室模型的真核细胞和生物体。原核生物缺乏细胞核的单细胞生物尚未在衰老研究中占据一席之地。在下一节“生物衰老学家如何研究衰老:比较生物衰老学”中,我们探讨了野生动物作为人类衰老和长寿模型的应用。
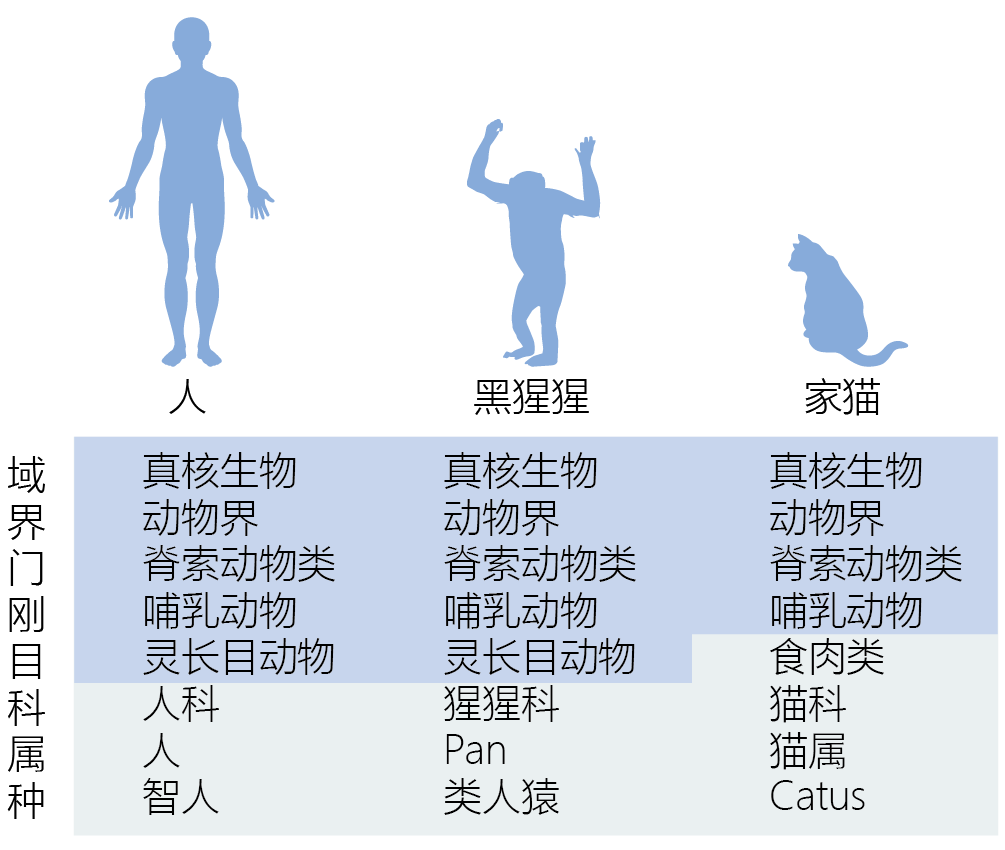
无论使用哪种生物,由于所有真核生物之间的系统发育关系,使得该研究将与人类衰老相关(表1.1)。系统发育学描述了基于基因相似性的生物体之间的相关性。
| BOX 1.1 生物系统树 |
|
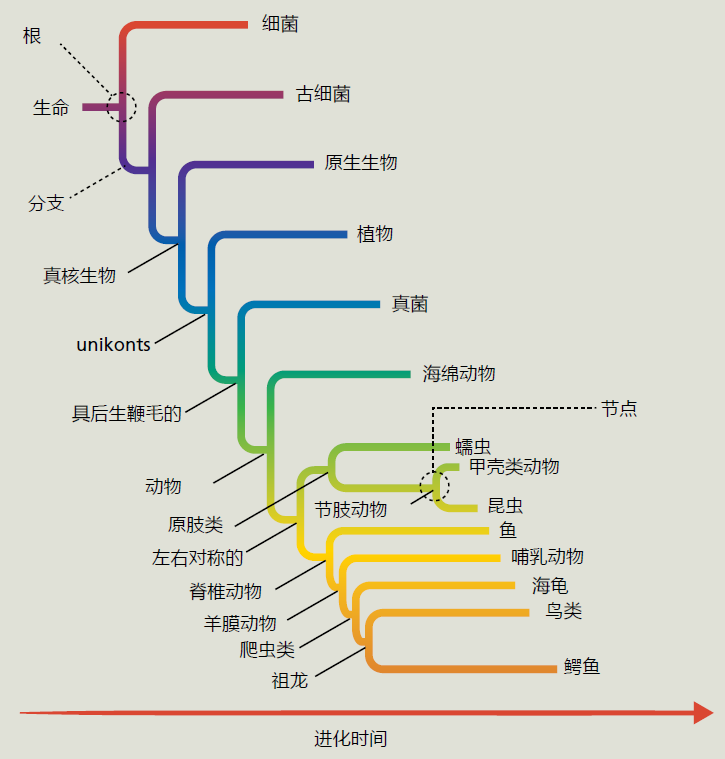
在二十世纪之前,生物多样性的分类及其相互关系受到几个世纪哲学和神学教义的极大影响。分类学的创始人约翰·雷(1627-1705)和卡洛斯·林奈(Carlos(Carl)Linnaeus)(1707-1778)发展了他们的生物体分类,以反映造物的神圣秩序,“秩序”是关键词。近200年来,Linnaeus的分类系统仅使用形态学(生物体的形态和结构)来表明进化朝着更复杂的方向前进——细菌是最简单和最早的生命形式,人类是最复杂和最新进化的生命形式(图1.6)。 图1.6林奈分类系统。Carolus Linnaeus的分类系统是一系列基于有机体与其他生命形式相似性的等级分类。虽然形态分类系统正被系统发育系统所取代,但林奈开发的分类名称仍被广泛使用。 在重新发现并最终理解孟德尔的遗传学原理之后,生物学家开始质疑进化是否真的反映了生命从低复杂性到高复杂性的有序过程。使用另一种分类形式的压力越来越大,因为发现所有生物体的DNA结构都是相同的,在“低等”生命形式中发现的许多基因与在“高等”动物中发现的基因是相同的。这些发现提供了确凿的证据,证明所有生命起源于一个共同的祖先,或者至多是几个共同的祖先。此外,进化论者发现形态的复杂性对于一个物种的进化史来说是一个糟糕的描述。物种的复杂程度取决于它们在环境中生存的需要。因此,物种的复杂性更多的是由于物种在其环境中生存的能力引起的,而不是随着系统进化树进行的。 二十世纪中后期生物科学的进步使得了基于系统发育而非形态学的分类系统的发展。系统发育是一个物种或一组有机体发展过程中所涉及事件的进化序列。现代系统发育学综合运用各种因素和技术来建立物种间的进化关系。这些技术包括形态特征、DNA序列、生态数据和可能的基因关系的数学算法。系统发育学并不认为一个物种比另一个物种更先进。它简单地认为,一个物种是从一个先前的群体进化而来的,这是由于对其近端环境的遗传适应。 系统发育关系可以使用系统发育树来呈现,这是一个分支图,显示了各种物种之间可能的进化关系(图1.7)。树的分支定义了单系群体之间的祖先和后代关系。单系群包含同一祖先的所有后代。 图1.7 来自同一个祖先、并且无标度的系统发育树。系统发育树由节点组成,每个节点代表一个分类单元(物种、种群、个体)和分支,分支根据血统和祖先定义分类单元之间的关系。例如,节肢动物是一个包含昆虫和甲壳类动物的单系类群。在系统发育树中,节肢动物由一个节点表示,昆虫和甲壳类动物由该节点的分支表示。 树的节点表示分类单位,如有机体、物种或由单个分支连接的种群。树的拓扑或分支模式可以缩放或不缩放。按比例生长的树使用与分类单元之间发生的进化变化数量成比例的分支长度。未缩放树仅使用分支连接关系。树木也可能有根或无根。有根的树,如图1.7所示,对于树上的所有其他物种或群体,都有一个共同的祖先。无根树只说明了关系,没有提及共同的祖先。 系统发育学不仅仅是一个分类系统,它还是一个有用的工具。例如,分子系统发育学家通过基因序列比较,从而在人类和家猪之间建立了密切的进化关系,表明我们和家猪在生理学上有着密切的关系。事实上,猪心在结构和功能上与人类心脏相似。医学人员利用这些信息来测试是否可以将猪的健康心脏瓣膜移植到衰竭的人类心脏中。事实证明,猪的瓣膜几乎与人类瓣膜完美匹配。今天,许多人活得很好,因为猪的心脏瓣膜替换了他们自己有缺陷的瓣膜,部分原因是系统发育学。 |
这里的讨论是对生物衰老学实验中普遍使用的物种的介绍。后面的章节将更详细地探讨这些模型系统的具体用途。这里不讨论植物;第六章将详细介绍植物生物衰老学。重要的是要记住,没有任何一种动物或植物模型是研究人类生物衰老的“完美”系统。实际上,我们选用哪种生物主要是看我们要研究哪些问题、该生物体的衰老和寿命、该生物体的繁殖类型和成功率,以及该生物体的护理和养护成本。
孤立的细胞系统可以用来描述衰老和长寿的基本生化过程
人类是地球上最复杂的生物之一,拥有复杂的神经、血管和内分泌系统,使我们在进化上取得了成功。尽管如此,为了有效地工作,这些先进的系统依赖于其细胞内生物化学的正常功能。因此细胞的功能以及细胞随时间的变化最终可以描述人类如何衰老。从细胞角度研究衰老在老年学中有着悠久的历史,可以追溯到1912年,当时第一次细胞培养成功(见第4章)。从这些早期的研究中产生了四种基本的细胞系统,它们被用于生物衰老的研究:原代细胞培养、复制细胞培养、细胞系和干细胞。
原代细胞培养是直接从其体内取出并在体外环境中培养的已分化细胞(图1.8)。在生物衰老学研究中,原代细胞培养通常含有有丝分裂后的细胞,或增殖能力有限的细胞,并且在很短的时间内保持存活,通常只有几天。原代细胞培养让研究人员能够比较一下特定类型的分化细胞之间的差异。例如,可以使用某些技术来确定平滑肌细胞的收缩特性。因此,年轻和老年动物的平滑肌细胞可以被移除,置于培养基中,然后评估与年龄相关的差异。
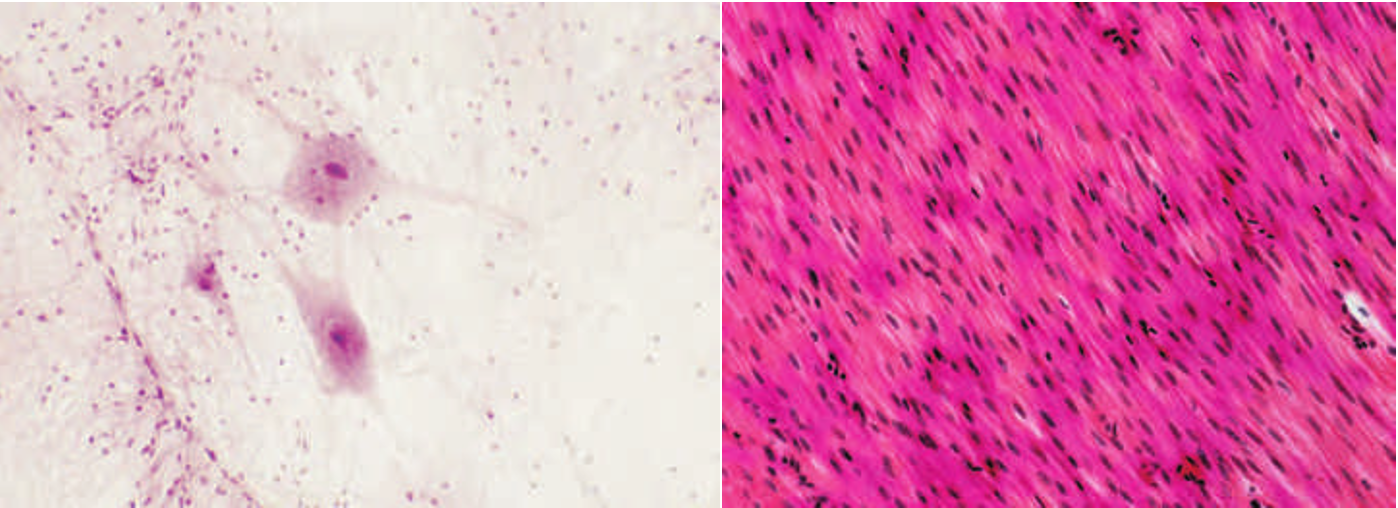
图1.8 原代细胞培养。(A) 人类神经元。(B) 平滑肌细胞。(A,由Thinkstock提供;B,由S.Gschmeissner/Getty Images提供。)
复制细胞培养是生物衰老学中应用最广泛的细胞培养系统。复制细胞培养是未分化的有丝分裂细胞,如成纤维细胞,它们已从组织中移除并允许分裂,直到达到接触抑制(培养皿范围内的最大容量)。然后将它们分散开并转移到另一个烧瓶中,让其再次生长,这一过程称为种群倍增。哺乳动物细胞可以在种群死亡前加倍30-50倍。通过在不同时间对培养液中的细胞取样,生物衰老学家可以比较年轻细胞和老年细胞中的细胞内的因子。这些系统通常用于评估导致细胞衰老和死亡的因素,因为有丝分裂细胞在体外具有有限的复制寿命。
细胞系是无限增值的有丝分裂细胞。这些细胞群要么来自癌性肿瘤,要么是变异了的正常细胞(其内部生物化学已经发生了改变,使其获得不死性)。细胞系是一般细胞生物学研究中的一个重要对象,但它们在衰老生物学中并未得到广泛应用,主要是因为这些细胞不会衰老,也不会表现出正常细胞中观察到的与衰老相关的功能丧失。然而,一些研究人员使用细胞系来研究衰老和癌症的共同途径(见第4章:细胞衰老可能保护细胞免受癌症)。干细胞是未分化的细胞,具有无限期自我更新的能力;它们分裂并产生一个分化的细胞。干细胞有两种形式,胚胎和成体。胚胎干细胞要么是全能的,具有生成整个有机体(包括胎盘)的能力,要么是多能干细胞,具有从三种胚层(内胚层、外胚层和中胚层)生成细胞和组织的能力(图1.9)。成体干细胞是多能的,从哪个组织中提取,就会形成哪个组织的细胞:肝干细胞产生肝细胞,肌肉干细胞产生肌肉细胞,等等。干细胞在生物衰老学研究中具有价值,因为它们具有使组织再生或替换组织的能力。例如,产生血细胞的造血干细胞在化疗后被植入老年患者的骨髓中,以通过加快再生过程降低感染风险。
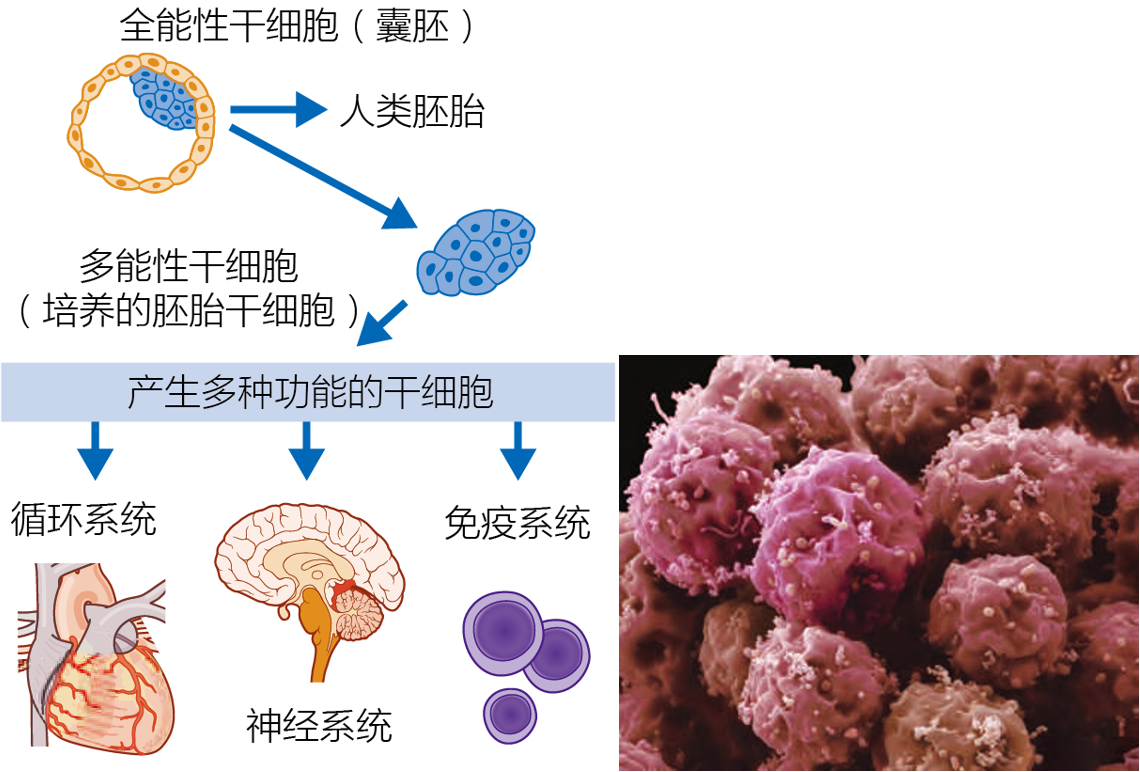
图1.9 胚胎干细胞可产生身体的不同类型细胞。(A) 胚胎干细胞(ES)从胚泡的内部细胞团中获取。这些细胞可以分化成特定的细胞类型。(B) 培养中的胚胎细胞克隆。由于其无限的自我更新能力,胚胎干细胞被认为是一种再生方式,替代哪些因衰老或年龄相关疾病而受损的组织和器官。(B,由S.Gschmeissner/科学图片库/盖蒂提供。)
如果不简要提及诱导多能干细胞(IPSC),我们关于干细胞在衰老研究中的应用的讨论将是不完整的。诱导多能干细胞是从基因重新编码的成人皮肤和血细胞中获得的干细胞。2006年,由诺贝尔奖获得者Laurate Shinya Yamanaka领导的一个日本研究小组报告说,他们成功地改变了成年小鼠皮肤细胞内的基因,并培养出与胚胎(多能干细胞)行为相似的细胞克隆。这些结果表明,现在有可能从我们自己的身体中提取皮肤或血细胞,并将它们转化为各种类型的细胞、神经、肌肉等。反过来,IPSC可以用来培育新的器官,取代那些因疾病和时间而受损的器官。使用从人类胚胎中提取的细胞将不再存在免疫排斥或伦理问题的风险。
通过IPSC进行组织置换的临床尚未在人类身上实现。截至本文撰写之时,只有一个人接受了来自IPCS的组织,这是一种眼部疾病的视网膜替代物。尽管移植后恢复了一部分患者的视力,但新组织的细胞发生了突变,这引起了操作该试验的眼科医生的安全担忧。没有在任何其他人身上进行进一步的试验。尽管如此,个体恢复视力的事实证明,用新的功能细胞替换受损或患病的组织是可能的。困扰新组织的意外突变问题以及与IPSC相关的其他问题将得到解决。总有一天,来自IPSC或类似IPSC的组织替代物将成为一种常见的外科治疗方法。
真菌是研究影响衰老和寿命的环境因素的良好模型
酵母和菌丝形式的真菌没有复杂的血管、神经或内分泌系统,使细胞间信号传递变得困难(细胞间的通讯通过细胞壁上的缝隙或孔进行)。它们必须依靠细胞与环境的直接接触来感知周围的世界。这一特性使真菌成为研究影响衰老的环境因素的极好对象。此外,真菌衰老的研究为研究人员提供了一些优势。首先,真菌在地球上几乎所有环境中都能生存(图1.10)。可以选择一种真菌,以符合研究人员假设的对衰老有影响的环境条件。其次,真菌的核基因组和线粒体基因组具有紧凑、较高的编码-调控序列比率。正如你在第5章中看到的,较高的编码与调控序列的比率使研究人员能够更精确地确定哪个基因具有哪个功能。第三,真菌的寿命范围很广,从几天到8000年不等。第四,大量的单个真菌可以在实验室以非常低的成本快速生长。

图1.10 真菌的多样性。真菌生活在不同的环境中,并且不同种类的真菌寿命跨度很大。(A) 出芽酵母(酿酒酵母)很容易在培养基中生长。(B) 蜜菇(蜜环菌;又称小花菌)可能是地球上最古老的生物。在俄勒冈州东北部的Malheur国家森林中发现的一株A.ostoyae可能有8000年的历史。(C)生长在热带雨林中的杯状真菌(Cookeina sulcipes)和(D)生长在冻土带上的驯鹿苔藓(Rangifera),都是能够在恶劣条件下生存的长寿真菌的例子。(A,由S.GSCHMEISNER/科学图片库/Corbis提供;B,由M.Watson/moodboard/Corbis提供;C,由M.Read/123RF提供;D,由A.Romanov/123RF提供。)
原始无脊椎动物可能为延长细胞寿命、细胞信号和全身衰老提供线索
原始无脊椎动物是一个多样化的群体,包括海绵、水母、海葵、珊瑚、蠕虫、轮虫和软体动物(图1.11)。许多水生无脊椎动物的寿命非常长,直到最近才在衰老研究中受到重视(见下一节,“生物衰老学家如何研究衰老:比较生物衰老学”)。

图1.11 长寿海葵。海葵,如这种巨大的绿色海葵(anthoplera sola),据报道具有极端的寿命和无限的生长。(由altrendo nature/Thinkstock提供。)
蠕虫和轮虫很容易在实验室饲养,而且大多数寿命相对较短。尽管与高阶动物相比,这些生物的细胞和组织特化是原始的,但这些物种通过细胞连接具有复杂的细胞间通讯。因为这些动物也有一个紧凑的基因组,它们是研究细胞事件如何与全身衰老联系的极好模型。第五章详细描述了C. elegans中生殖开始时与环境相关的细胞信号通路的遗传操作,从而发现可能调节寿命的基因。此外,这些生物中有许多是真生物;也就是说,它们成熟时有固定数量的细胞。由于这些物种中有一些无法更新组织,这些物种可能会为生物衰老学家提供一个研究随机衰老的模型,这是你在第3章和第4章中了解的全身衰老原理。
昆虫可以用来研究全身和细胞内信号如何影响生命史
昆虫是地球上最大的一类动物,已知物种有300万种,未发现物种的数量是已知物种的数倍。许多昆虫的寿命短,繁殖率极高,这为在短时间内研究和操纵几代昆虫的遗传学提供了机会。此外,昆虫的生活史(生物体在其整个生命周期内发生的所有生物事件的总和)比许多其他更复杂的动物的生活史更容易通过环境操纵来调节。例如,昆虫的繁殖活动,以及它们的寿命,可以通过温度、食物供应和白天光线的变化来改变。这种类型的调节通常与神经内分泌系统信号的变化有关。因此,研究人员可以利用昆虫来研究全身和细胞内信号如何影响生物体的生活史。
虽然昆虫作为人类衰老模型的好处是显而易见的,但只有少数物种在很大程度上得到了研究。果蝇,黑腹果蝇,被广泛用于衰老研究,是第一种精确测定其寿命的动物。D.melanogaster在衰老研究中的定位主要在于长寿遗传学的研究,并将在后续章节中详细讨论。
小鼠(mouse)和大鼠(rat)是研究营养、遗传和生理问题的常见研究对象
绝大多数生物衰老学研究都是以大鼠或小鼠作为模式生物进行的。啮齿动物在研究中特别有用,因为啮齿动物与人类生理学和细胞功能相似。与其他寿命相似的动物相比,小鼠和大鼠的饲养成本相对较低。与人类受试者不同,啮齿动物的饮食和环境可以严格控制。此外,啮齿类动物可以很容易地进行基因操作,从而可以测试基因产物和与年龄相关的变化。由于目前的许多研究都是在这些动物身上进行的,它们在研究中的具体用途将在后面的章节中详细描述。
非人灵长类动物表现出许多与人类相同的时间依赖性变化
非人灵长类动物在基因上与人类最接近,因此是研究人类衰老生物学基础的最终模型。几种非人灵长类动物,如狐猴、狨猴、猴子和大猩猩,已被用于研究衰老生物学,但大多数控制良好的实验室研究涉及恒河猴。衰老的恒河猴表现出许多与时间相关的生理衰退,这些生理衰退也在人类身上观察到,而在其他物种中并不经常观察到(图1.12)。这些衰退包括视觉和听觉缺陷、运动功能下降、骨矿物质含量下降、雌性真正绝经和雄性睾酮水平下降、肌肉质量下降以及代谢功能普遍下降。

图1.12 恒河猴。非人类灵长类动物,如恒河猴,由于其与智人的遗传相似性,是人类衰老的宝贵模型。如图所示,恒河猴显示出明显的衰老迹象:(A)一只18个月大的猴子;(B) 一个25年的猴子。他们还易患许多与年龄有关的疾病和人类常见的功能障碍。(A,由J.Lenon提供;B,由J.Miller提供。)
恒河猴也易患许多依赖时间的人类疾病,如2型糖尿病、心血管疾病以及阿尔茨海默病和帕金森病的伪型。在恒河猴和人类中,2型糖尿病和心血管疾病的病因似乎是相同的。
非人灵长类动物和人类在时间依赖性功能丧失和疾病方面的相似性为科学家提供了可以严格控制的群体,在这些群体中,科学家可以重复进行非侵入性或低风险侵入性程序,测试药物和其他物理疗法。例如,许多目前可用于人类的骨质疏松症和抗骨质流失处方药都在恒河猴身上进行了试验。然而,使这些物种作为衰老模型具有吸引力的相同生理相似性也存在主要局限性。与人类的基因相似性导致了对这些近亲进行侵入性实验的伦理问题。负责确保研究动物得到人道待遇的监管机构做出了回应,限制了可以在非人灵长类动物身上进行的研究类型。一般来说,非人灵长类的研究方式仅限于人类也允许的方式,尽管安全标准可能较低(例如,猴子的X射线照射限值高于人类)。
使用非人灵长类动物作为人类衰老模型的另一个主要限制是成本。动物必须在其整个生命周期内保持在高度受控的条件下。居住在认证动物设施中的恒河猴的平均寿命约为35年,最长寿命接近45年。养一只恒河猴的平均费用因研究机构而异,但每天费用一般在15至20美元之间。因此,养一只恒河猴35年的费用高达25.55万美元。个体研究者不太可能获得足够的资金,以完成一项良好控制的老龄化研究所需的恒河猴数量进行调查。例如,国家卫生研究院目前仅在全国三个地点支持对恒河猴的研究。
人类早衰可以用来模拟正常的人类衰老
Werner综合征和Hutchinson-Gilford早衰综合征是许多人认为与早衰有关的疾病(图1.13)。早衰是一种罕见的遗传病,其特征是身体生长缓慢,并具有快速衰老的特征性迹象。Hutchinson-Gilford早衰综合征在出生时和幼年时影响患者,而Werner综合征通常在患者的第二或第三个十年开始表现出来。虽然这两种综合征都会增加个体罹患年龄相关疾病的风险,但Werner综合征患者往往死于癌症和动脉粥样硬化,而Hutchinson-Gilford早衰综合征患者更容易患心血管和神经系统疾病。

图1.13 患有Werner综合征的妇女。图中所示为同一个人,分别是13岁(A图)和56岁(B图)。(A,摘自Hisama FM et al.2006.Sci Aging Knowl Environ 10:18.经美国科学进步协会许可;B,由J. Oshima提供。)
因为Werner综合征患者的寿命往往比Hutchinson-Gilford早衰综合征患者的寿命更长(45-50岁与12-15岁),Werner综合征被认为是更好的衰老模型。Werner综合征患者共有四个特征:身材矮小、头发早期变灰并脱发、双眼白内障和硬皮病样皮肤改变。许多患者还有扁平足、声音改变和性腺机能减退。Werner综合征患者患2型糖尿病、动脉粥样硬化、冠心病、高血压和骨质疏松症的风险很高。这些外观上的变化和与年龄有关的疾病风险的增加显然与正常衰老过程中发生的变化相似。
Werner综合征是由wrn基因突变引起的。该基因负责产生WRN蛋白,该蛋白在DNA的维持和修复中发挥作用。这种蛋白质也可能有助于DNA复制。WRN蛋白功能的缺失或降低与第4章讨论的一些衰老理论预测的结果相似。