2.先天免疫系统
一直以来,大多数免疫学家都不十分重视先天性免疫系统,也许因为获得性免疫系统更为令人兴奋,然而,对获得性免疫系统的也就已经使先天性免疫系统的作用得到了新的认识,其作用不仅是快速反应的第二屏障,而且还是获得性免疫系统的激活剂和控制者。
如果设想一下细菌感染不受控制时可能发生的事情,就不难理解先天性免疫系统对普通入侵者作出快速反应的重要性了。假设你的浴盘碎片刚好把一个细菌带入你的组织。正如你知道的,细菌的繁殖是非常快的。实际上,单个细菌每30分钟倍增,在一天内就能繁殖大约100万亿个细菌。如果你培养过细菌,你就知道含有1万亿个细菌的1升培养基是如此稠密,以至于你的视线不能看穿它。因此单个细胞增殖1天就能产生约 100升的稠密培养物。然而不要忘记你的血液总量大约只有5升,你就可以理解未受抑制的细菌感染对人所能造成的危害了。如果没有快速反应的先天免疫系统包括补体蛋白,我们将会面临很大的麻烦。
先天性免疫系统包括补体蛋白,专职吞噬细胞和自然杀伤细胞。我们将从补体系统开始讨论。
2.1 补体系统
补体系统由大约20种不同蛋白质造成,这些蛋白质通过共同作用来消灭入侵者并把信号传递给参与进攻的其他免疫系统成员。补体系统是一个非常古老的免疫系统,即便是进化了约7亿年的海胆都已经具有了补体系统,对人类而言,在胎儿发育的前 3 个月补体蛋白就开始合成了,因此,很显然在孩子出生前机体就已经准备好了这个重要系统。事实上,很少有婴儿生下来就有补体蛋白的缺陷,而补体蛋白缺陷的孩子在死于感染前就很难存活。
当我第一次接触到补体系统时,我认为它很复杂,难以理解。但当我深入研究时,我开始认识到,它其实是非常简单,也是如此美丽。就像免疫系统种的其他组分一样,补体系统在发挥作用前必须激活,补体激活的途径有3条。第一条是所谓的经典途径,依赖于抗体的激活,这一途径我们将在后面讨论。补体系统作用的方式不依赖于其激活的途径,因此其后讨论抗体依赖的激活途径不会影响你对其他系统的理解。
补体激活的第二条途径称为旁路途径,虽然作为一个进化学术语,旁路途径源于经典途径之前,但免疫学家还是把抗体依赖的系统称为经典,仅仅因为它是第一个被发现的。直到最近,免疫学家还认为这是补体系统能被激活的仅有的两条途径。然而,现在已经发现了第三条途径,即凝集素激活途径。
2.1.1 旁路途径
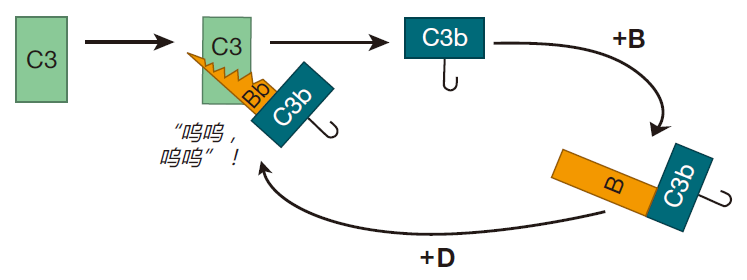
构成补体系统的蛋白质主要由肝脏合成,而在血液和组织种存在的浓度较高。最丰富的补体蛋白是 C3,在人体内,C3 分子可被连续降解成两个较小分子的蛋白质。由这种自发切割产生的蛋白质片段之一的 C3b,是非常有反应性的,并能和两种常见的化学基团(氨基或羟基)结合。由于许多构成入侵者表面(如细菌的细胞壁)的蛋白质和糖类都有氨基或羟基,因此这些小 C3b 就有许多靶分子。
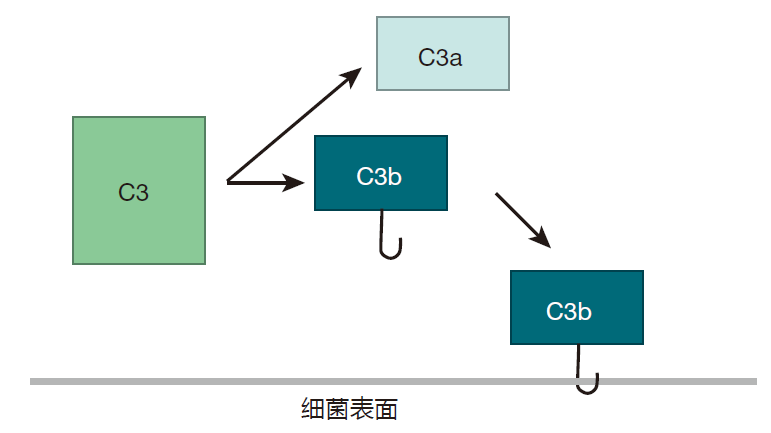
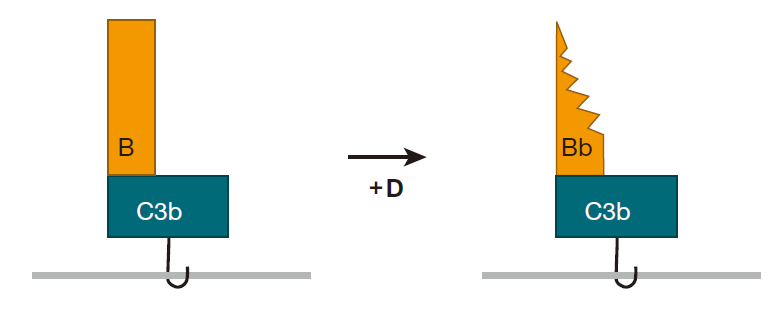
如果 C3b 在大约 60 微秒内不能和这些化学基团种的一种发生反应,它就通过和一个水分子结合而被中和,“游戏”也就因此而结束。因此,为了能使补体连锁反应得以持续,自发裂解C3分子必须接近细胞表面。一旦C3b稳定结合于细胞表面,另一个补体蛋白——B就裂解到C3b上,随后补体蛋白D参与反应并切下一部分B,从而产生 C3Bb。
一旦细菌被这个C3Bb 分子粘住其表面,有趣的事就真正开始了,因为 C3Bb的作用就类似一条“链锯”,能切开其他 C3 蛋白并把 C3 蛋白转变成 C3b。这样邻近的 C3 分子无需自发裂解而即可形成 C3b,因为 C3Bb 分子(称为“转化酶”)能非常有效地行使上述职责。并且一旦其他的 C3 分子被裂解,其产生的 C3b也能和细菌表面的氨基或羟基结合。
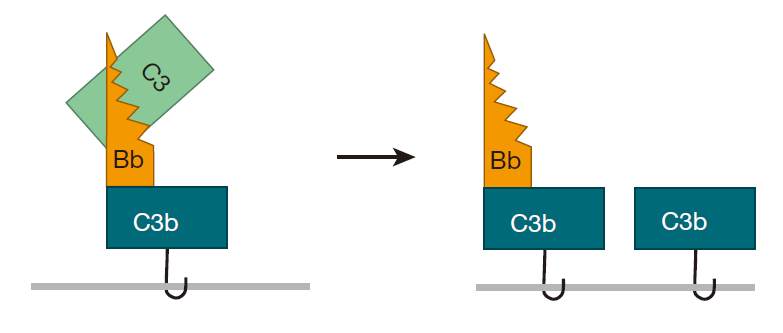
这个过程一直持续,不久就会有大量 C3b 分子粘附到靶细胞表面,而每个C3b 都能形成一个 C3bBb 转化酶,从而能切开更多的 C3 分子。正如你看到的,所有这些粘附和切割形成了一个正反馈环路,并且整个过程就像滚雪球般扩大。
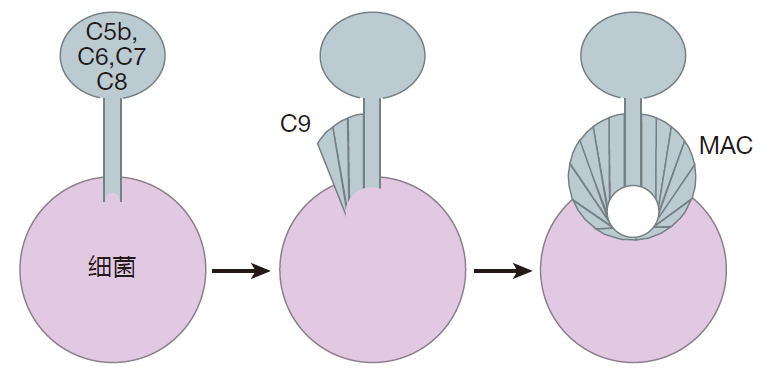
一旦 C3b 结合至细菌表面,补体级联反应就能进一步进行。C3bBb 链锯也能切开另一个补体蛋白——C5 的一部分,切割产物 C5b 能和其他补体蛋白(C6,C7,C8 和 C9)结合形成膜攻击复合物(MAC)。为了形成这种结构,C6,C7,C8 和C9 要形成一个“”主杆(stalk),把复合物锚定在细菌细胞壁上。然而C9蛋白加入该复合物形成一个通路,在细菌表面打开一个孔。一旦细菌表面形成了孔,它的噩运就开始了!
现在,你可能会感到疑惑:既然这些手榴弹会遍地开花,为什么补体系统不在我们自己的细胞表面形成 MAC?答案是因为人体的细胞装备了许多防护设施防止这种情况发生。实际上,机体是如此担心补体系统的异常反应,以至于她不得不通过与补体系统自身具有的蛋白等量的蛋白质来控制补体系统。例如,在人体的细胞表面有一种被称为衰变加速因子(DAF)的蛋白质,可以和血液中其他蛋白质一起作用,加快转化酶 C3Bb 的分解作用。补体片段 C3b 也能被血液中的蛋白质修剪成一个无活性的结构,存在于人体细胞表面的酶也可以加速这种修剪作用,其他细胞表面蛋白如 CD59(也称保护素,protectin)在 MAC 钻孔之前就能把几乎形成的 MAC 全部“踢开”。
我们可以用一个有趣的故事来阐明这些保护设施为什么如此重要。如你所知,移植外科医生没有足够的人体器官来满足移植需求,因此他们正考虑使用动物器官,其中一个热门的器官供体候选者是猪,这是因为猪饲养价廉,其器官和人的器官大小相近,随着人体移植的升温,外科医生决定把猪的器官移植到狒狒体内,结果并不成功,几乎是立即,狒狒的免疫系统开始攻击器官,并且几分钟之内移植的器官变成了血浆。元凶是谁呢?补体系统。原来,来源于猪器官的DAF和CD59不能控制灵长类的补体系统,因此,不受保护的猪器官易于受到狒狒补体系统的攻击。
这个故事强调了补体系统的 2 个重要特征,第一,补体系统的作用非常快。
补体蛋白以高浓度存在于血液和组织中,它们时刻装备抗御表面带有多余羟基或氨基的入侵者。第二,如果细胞表面没有受到保护,它将受到补体的攻击。实际上,你应该有这样一个印象,即补体系统不断投下这些小的手榴弹,如何没有保护的表面都将是其目标。在这个系统内,缺少选择权就是死亡。
2.1.2 凝集素激活途径
除了经典和旁路的补体激活途径外,还有最近发现的第三条途径,它可能是最重要的一条激活途径。这条途径的核心成分是一种主要在肝脏中合成的蛋白质,其在组织和血液中的浓度为中等。这种蛋白质称为甘露糖结合凝集素(MBL)。
凝集素是一种能和糖类分子结合的蛋白质,甘露糖则是一种常见于许多细菌表面的糖类分子。MBL 可以结合于酵母菌如白色念珠菌,病毒如 HIV 和 A 型流感病毒,许多细菌包括沙门菌和链球菌以及寄生虫如利什曼原虫等。与此相反,MBL却不能与健康人体细胞和组织中的糖类结合。
MBL 激活补体系统的工作方式非常简单。在血液中,MBL 与另一种称为 MBL相关丝氨酸蛋白酶(MASP)的蛋白质结合,然后,当 MBL 捕获到目标(如细菌表面的甘露糖)时,MASP 就发挥转化酶的功能,切开补体蛋白C3,从而产生C3b,由于C3b在血液中含量最丰富,因此这种作用非常有效。随后,C3b片段能和细菌的表面结合,从而启动我们刚才讨论的补体连锁反应并使其运行。
如上,旁路激活途径是自发的,并可以设想手榴弹随机地到处爆炸,用于破坏未受保护的表面;而凝集素激活途径可以被认为是一种灵敏炸弹,通过MBL来确定目标。这是先天性免疫系统所应用的主要策略的一个范例:先天性免疫系统主要针对于常见病原体表面的糖类和脂类分子。
2.1.3 补体系统的其他功能
除了建立MAC,在先天性免疫中补体系统还有其他两个功能。当C3b 粘附到入侵者表面时,它能被血清蛋白切割产生一个较小片段——iC3b,i 表示这个切开的蛋白多余生成 MAC来说是没有活性的。然而,它仍然可以粘附入侵者,以类似于抗体调理入侵者的方式为其后的吞噬作用加工处理入侵者。在吞噬细胞如巨噬细胞表面有补体受体能结合iC3b,iC3b调理入侵者结合有利于吞噬作用,这是一个重要的功能,由于许多入侵者都有相当粘滑的表面,使它们难以被巨噬细胞捕获。当这些粘滑的入侵者被补体片段包住时,吞噬细胞能更好地捕捉。因此补体的第二个功能就是修饰入侵者表面。
补体系统还有第三个重要功能:补体蛋白片段能充当化学趋化剂——即把免疫系统的其他成员吸引到战场的化学物质。例如C3a和C5a是C3和C5的裂解片段,C3和C5裂解时还生成C3b和C5b,这些片段不和入侵者的表面结合,而是处于自由状态,在它们发挥作用的组织中吸引巨噬细胞和中性粒细胞并激活这些细胞从而使它们成为更有潜力的杀伤细胞。有趣的是,C3a和C5a这类片段被称为过敏毒素,这是因为它们能促进过敏性休克,有关这部分的内容我们将在以后的章节中讨论。
因此,补体系统是一种多功能的系统,它可以通过形成 MAC 消灭入侵者,通过激活吞噬细胞上的补体受体而加强其功能,并且还能向其他细胞发出信号使攻击得以继续。最重要的是,它能非常快速地完成所有这些事情。
2.2 专职吞噬细胞
专职吞噬细胞是先天性免疫系统的第二件武器,这些细胞被称为专职细胞是由于它们主要通过吞噬作用来获得生存,最重要的这是吞噬细胞是巨噬细胞和中性粒细胞。
2.2.1 巨噬细胞
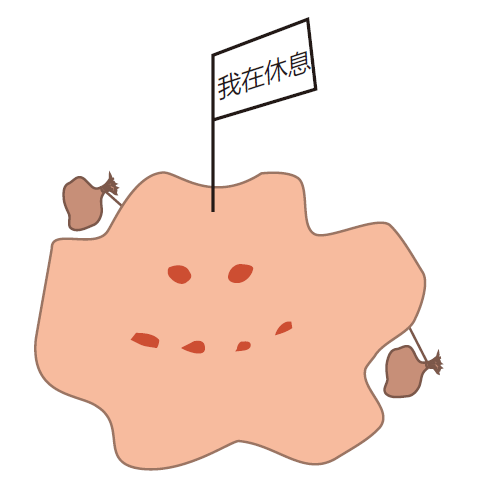
巨噬细胞可能有三种准备阶段。在组织中,巨噬细胞通常是闲荡的,而且增殖缓慢。在这个静息阶段,它们的主要功能是作为清道夫,无论周围有什么都要一点一点吞掉,以此保持我们组织中没有废渣。静息时,它们的表面只表达非常少的MHCⅡ类分子,因此它们不是很善于把抗原提呈给辅助T细胞,这相当有意义。为什么它们要收集垃圾呢?对正常的巨噬细胞而言,生命是相当枯燥的。
它们在组织中存活几个月,就要收集几个月的垃圾。
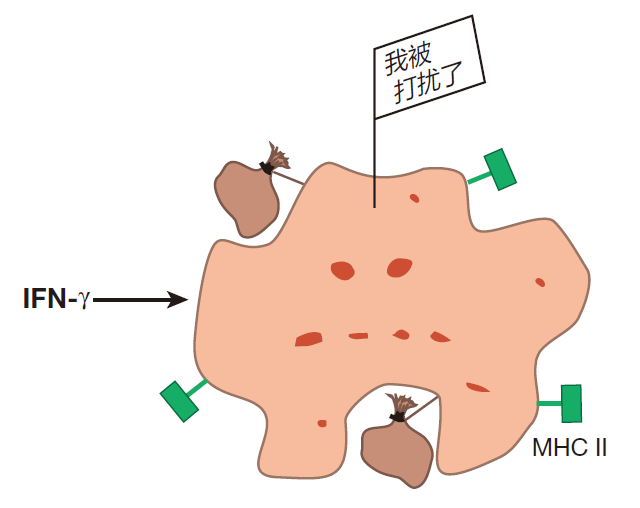
然而,一些静息的巨噬细胞偶尔也会收到让它处于待命状态的信号,即防御屏障被突破以及有入侵者攻入该区域。当这种情况发生时,它们就被激活(免疫学家称为“致敏”),在这个状态,巨噬细胞的吞噬功能加强,MHCⅡ类分子的表达也上调。这样,被激活的巨噬细胞能发挥抗原提呈细胞的作用,当它们吞食入侵者时,它们可以通过 MHCⅡ类分子来把入侵者蛋白的片段展示给辅助T细胞看。尽管许多不同的信号都能致敏静息的巨噬细胞,但研究最清楚的是 IFN-γ,它主要由辅助T细胞和自然杀伤细胞产生。
在致敏状态,巨噬细胞是很好的抗原提呈者和杀伤者。然而,它们还可以加入一个更高的准备状态——“超活化”状态。例如脂多糖分子就可以传递这种信号。LPS是革兰氏阴性菌如大肠杆菌的细胞壁外层的一种组分,可由这些细菌分泌,并结合到致敏巨噬细胞表面的受体上,巨噬细胞也有甘露糖的受体,如前所述,甘露糖是许多常见病原体细胞壁的糖类组成成分,也是一种能够激活补体系统的“危险信号”。当巨噬细胞表面的受体与 LPS 或甘露糖结合时,巨噬细胞就确知有入侵者,一旦意识到此,巨噬细胞就停止增殖,并把注意力转向杀灭。在这个超活化状态,巨噬细胞体积变大,吞噬效率也增高。实际上,巨噬细胞变得如此之大和有吞噬力是为了吞摄那些像单细胞寄生虫一样大的入侵者。当被超活化,巨噬细胞就可以产生和分泌 TNF,这种细胞因子能杀死肿瘤细胞和病毒感染的细胞,也能辅助激活免疫系统细胞。
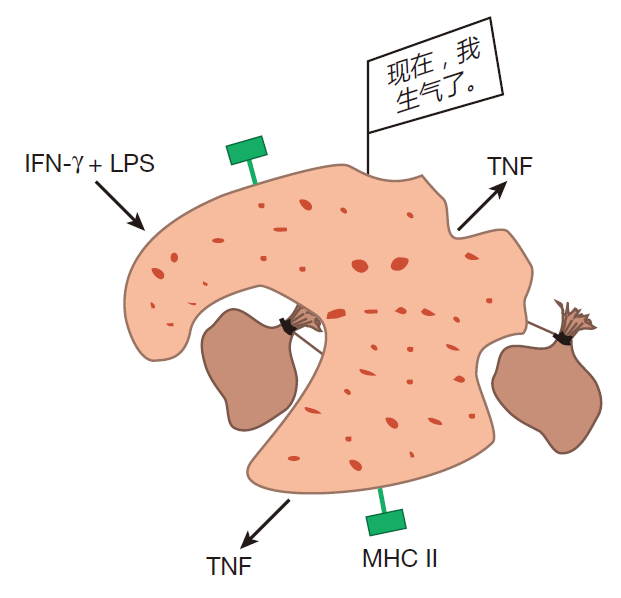
在超活化的巨噬细胞内,溶酶体数量增加,因此可以更有效地杀死吞入的入侵者。另外,超活化的巨噬细胞内反应性氧分子如过氧化氢的产生也会增加。过氧化物可以用来烫发,因此你可以想象它会怎么样对付细菌了,最终,超活化的巨噬细胞可以使其溶酶体的内容物作用于多细胞寄生虫,从而使它能消灭那些因太大而不能“吞食”的入侵者。是的,超活化的巨噬细胞就如同一部杀戮机器。
巨噬细胞是一个多面手,依据它的不同的激活阶段,既可以作为清道夫、APC,又可以作为凶恶的杀手。但是,你不应该形成这样一种印象,即巨噬细胞有三个“变速器”。免疫学中没有如何东西有变速器,巨噬细胞的激活状态是依赖于所接受到的激活信号的类型和强度而保持的一种持续状态。
2.2.2 中性粒细胞
尽管在功能多样性方面巨噬细胞是无可比拟的,但最重要的专职吞噬细胞可能还是中性粒细胞。中性粒细胞约占血液循环中白细胞的70%,在我们的骨髓中每天约产生1000亿个这种细胞,显然它们非常重要,否则我们就无需产生如此多的中性粒细胞了。中性粒细胞生存的时间非常短暂。实际上,它们从骨髓中产生出来以后平均在5天之内就会程序性走向死亡 。有趣的是,它们通过一种被称为凋亡的方式进行自杀。和巨噬细胞相反,中性粒细胞不是APC,它们是专职杀伤细胞。当中性粒细胞离开血液时,它们就被激活了,在这种状态下,它们非常类似于超活化的巨噬细胞,具有惊人的吞噬能力,一旦它们的猎物被吞入,一组强有力的化学物质就等待这个不幸的“客人”了。
我的朋友Dan Tenen研究中性粒细胞,他的妻子Linda Clayton从事T细胞实验,喜欢嘲弄他“为什么这么枯燥地研究中性粒细胞呢?它们所做的只是跳进脓液并走向死亡!”她是对的,脓液主要就是由死亡的中性粒细胞组成。然而,Dan提醒她说,没有那些奇妙的T细胞,人类能生存很长时间,但如果没有了中性粒细胞,那他们将在几天之内因感染而死亡。
那么,你认为是什么原因促使机体可以让巨噬细胞存活很长时间,而中性粒细胞却只能存活几天?这不是浪费吗?为什么不让中性粒细胞也像巨噬细胞一样拥有长的寿命?如果真是这样,那就太危险了,中性粒细胞从血管出来就是准备杀戮,在这个杀戮过程中,对正常组织而言,总是有危险存在的,因此为了减少危险,中性粒细胞就被只能有短的生命期。如果战斗需要更多的中性粒细胞,可以从血液中得到补充,那里总是有大量的中性粒细胞,相反,巨噬细胞却必须存活很长时间,因为它们就像哨兵一样,监视入侵者并发出攻击信号。
你可能感到疑惑:如果中性粒细胞真的那样危险,那么它们是如何知道何时应该离开血液,又前往何处呢?中性粒细胞肯定不会在原处离开血液并被激活。的确不会,这一过程目前已经非常清楚。在血管内部,中性粒细胞以非活性的状态存在,它们在血液中一很高的速率穿行,大约每秒 1000微米。如果你和中性粒细胞一样大,你就会知道这是多么快了。
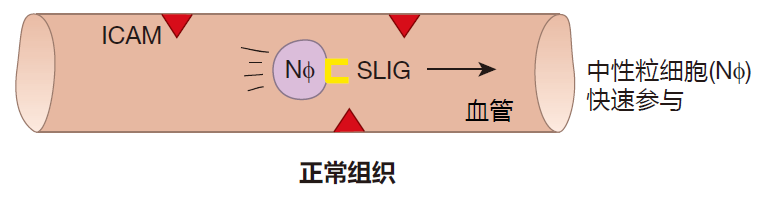
在上图中,你会注意到有一种蛋白质 ICAM,这种蛋白质表达于血管内皮细胞的表面,而在中性粒细胞表面则有另一种称为选择素配体的黏附分子表达,然而,正如你看到的,这两种黏附分子不是“配偶体”,因此,它们不会相互结合,中性粒细胞可以和血液一起自由穿行。
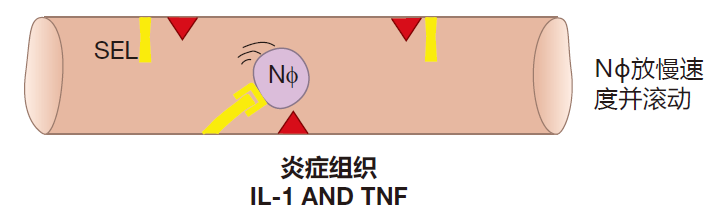
现在,假设你的大脚趾被一个木屑划伤了,碎片上的细菌激活了脚组织中担任警卫的巨噬细胞,这些被激活的巨噬细胞释放“警报”细胞因子——IL-1和TNF,从而发出了这里有入侵的信号,当邻近的血管内皮细胞收到这些警报信号后,它们开始在表面表达一种称为选择素的新蛋白质,合成这种蛋白质并运输到血管内皮细胞表面通常需要约6个小时。选择素是选择素配体的黏附伴侣,因此当选择素在血管内皮细胞表面表达时,它可捕获从旁边急驰而过的中性粒细胞。然而,选择素和配体之间的相互作用只有足够的强度才让中性粒细胞慢下来,并沿着血管内表面滚动漂流。
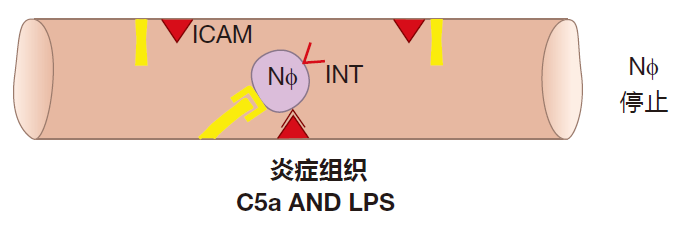
在中性粒细胞漂流时,它能缓慢地嗅察组织中有无“战斗”(炎症反应)进行的信号。补体片段C5a和细菌细胞壁成分LPS是中性粒细胞可以识别的两种炎症信号。一旦它收到这种信号,中性粒细胞就可以快速发送一种新蛋白——整合素至其表面。这种快速反应非常重要,由于中性粒细胞还没有停下来,仍然在漂流。如果它漂流的太远了,它将会离开选择素表达的区域,并再次提速到“血流速度”。为了尽可能地得到快速表达于表面的整合素,中性粒细胞会预先合成许多这种蛋白质,并储存在细胞内直到需要时使用。
当整合素出现在中性粒细胞表面时,就与表达在内皮细胞表面的ICAM 发生相互作用,这种相互作用非常强烈,它可以使中性粒细胞停止漂流。
一旦中性粒细胞停下来,它就受到一种称为化学趋化因子的影响,避开血管内皮细胞的监视,进入组织并迁移到炎症的位置,这些化学趋化因子包括我们已知的老朋友——来自于补体系统的C5a以及称为f-met肽的细菌蛋白片段。
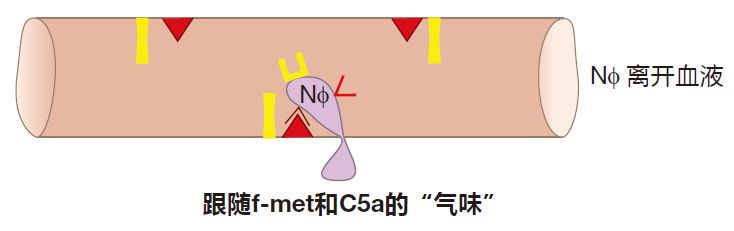
所有细菌蛋白都有一个特别的起始氨基酸,这种氨基酸称作为甲酰甲硫氨基酸(f-met),而在人体蛋白中 f-met的含量则少于0.1%,因此 f-met肽对细菌而言是相对独特的。当巨噬细胞摄入细菌时,它会吐出 f-met肽,中性粒细胞离开血液后就可以跟随这些f-met肽找到战场。另外,细胞因子如TNF也会激活在组织中穿行的中性粒细胞,使它们到达战场开始杀戮。
这一涉及了选择素-选择素配体的结合以使中性粒细胞漂流,整合素-ICAM相互作用使中性粒细胞停止漂流,化学趋化因子和中性粒细胞上的受体结合促进中性粒细胞离开血液的系统看起来似乎过于复杂。如果让一对黏附分子(如选择素及其配体)做这三件事是否会更简单呢?是的,这可能会更简单,但也可能非常危险。人体中大约有1000亿个内皮细胞,如果它们之中的一个细胞有一点异常,并开始在表面表达大量的选择素。如果选择素的结合是唯一的条件,那么,中性粒细胞能从血液全部流入正常组织,在那儿造成巨大的破坏。因此,在中性粒细胞离开血液之前,必须表达三种分子使得这个系统更为安全。
你应该还记得我曾说过第一种细胞黏附分子——选择素的完全上调表达需要大约6个小时,这是不是有点太慢了呢?一旦巨噬细胞感觉到危险,就开始从血液中补充中性粒细胞,是不是更好吗?不一定。在开始补充援军之前,首先必须确定攻击非常严重,如果巨噬细胞只遇到少数入侵者,它通常在短时间内不需要帮助就能处理。相反,涉及许多巨噬细胞在内的严重入侵却能持续数天。参与战斗的许多巨噬细胞持续表达警报细胞因子多余选择素的上调表达是必需的,这样可以确保只要真正需要时才会有更多的部队被召集。
我想你们应该注意到这个系统的第二个特征是,尽管中性粒细胞占血液中白细胞总数的70%左右,但在血管外的正常组织中性粒细胞却很少。中性粒细胞是随叫随到的细胞,那么谁来完成这一使命呢?是哨兵细胞——巨噬细胞。我们拥有了一个伟大的防御策略,在这一策略中,当需要帮助时清道夫就会使“雇佣兵”警觉。正是巨噬细胞和中性粒细胞之间的这种合作关系使得整个系统得以工作。
中性粒细胞并不是需要离开血液进入组织的唯一的血细胞。例如,参与防御寄生虫感染的嗜酸性粒细胞和肥大细胞,必须离开血液到达寄生虫感染的部位,要想最终称为组织中的巨噬细胞,单核细胞也需要在适当的地方离开血液。另外T细胞和B细胞也必须离开血液进入淋巴结,在那儿它们才能被激活。并且一旦被激活,T细胞和B细胞就必须很快分散至感染部位。整个过程就像一个邮递系统,在这个系统中有几万亿个包裹即免疫系统细胞需要被投递到正确的目的地。
这个投递问题可以通过使用和中性粒细胞相同的一个基本策略来得到很好的解决。免疫系统“邮政服务”的关键特征是,引起细胞漂移和停止的分子随细胞类型和目的地的不同而不同。结果,这些细胞黏附分子的作用类似于“划分邮政编码”,以确保细胞被投递到适当的位置,如你所知,选择素分子及其配体都归属于各自的一大家族,只有选择素家庭的特定成员会和选择素配体家族特定成员配对。整合素及其配体的情况也类似。由于有这种两位数字的邮政编码(选择素类型,整合素类型),使得有足够多的“地址”可以用于把许多不同免疫系统细胞发送到正确的位置。通过给免疫系统细胞配备不同的黏附分子,以及给它们的目标配备相应的黏附配体,机体就可以保证不同类型的免疫系统细胞可以准确地漂移、停止并在合适的位置离开血液。
2.3 自然杀伤细胞
自然杀伤细胞(NK)最初被称为大颗粒淋巴细胞,因为类似于专职吞噬细胞,它们的胞内充满了包含化学物质和酶的颗粒。和其他血细胞一样,NK 细胞也是从干细胞分化而来的,并可和淋巴细胞(T 细胞和B细胞)分在同一个家族。
和中性粒细胞相同,NK 细胞也运用“漂移、停止、离开”这一策略来离开血液并进入感染部位的组织。只要在组织中,NK 细胞就十分有用,它们能杀伤肿瘤细胞、病毒感染细胞、细菌、寄生虫和真菌。有趣的是,NK 细胞通过促使被感染细胞自杀来行使其杀伤功能。有时,NK 细胞通过穿孔素蛋白把“自杀”酶如颗粒酶B运入靶细胞,另一种情况是,NK 细胞表面的 FasL 和靶细胞表面的 Fas相互作用,从而起始靶细胞走向自杀的信号传递。
NK 细胞怎样识别应该杀伤的细胞仍是一个谜。它们识别靶细胞的方法和杀伤性T细胞非常不同,或者通过它们的TCR来识别入侵者的小片段,NK细胞没有TCR,它们必须识别由 MHC蛋白爱提呈的除多肽之外的其他一些东西。最新的研究认为,NK细胞通过两种信号识别靶细胞:“杀伤”信号和“不杀伤”信号。
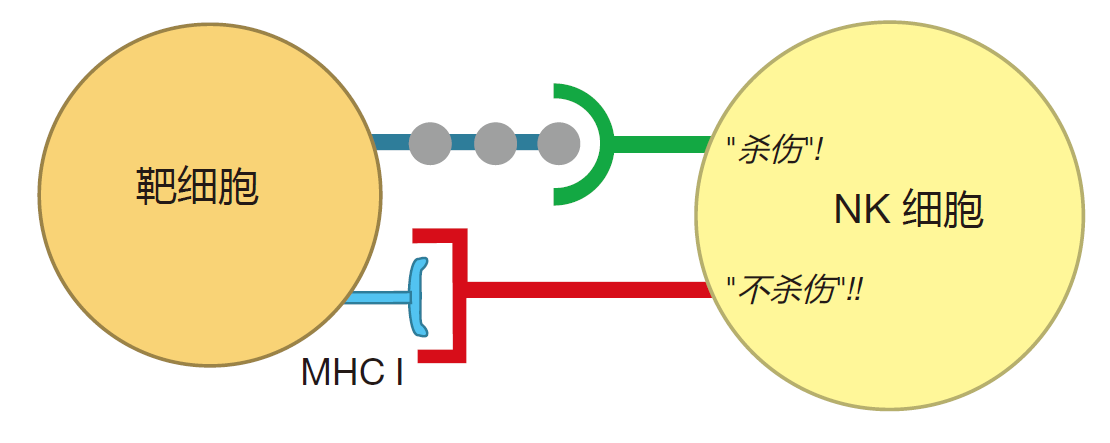
“不杀伤”信号是由在潜在靶细胞表面表达的分子来传递。研究最透彻的这类“抑制分子”是MHCⅠ类分子。表达MHCⅠ类分子的细胞通常不会被NK细胞杀死。“杀伤”信号通常被认为是由NK细胞的表面蛋白和靶细胞表面的特异性糖类分子或蛋白分子之间的相互作用来传递的。这些不平常的表面分子可能类似于一面旗子,表明细胞受到病毒感染或已成为肿瘤细胞,我们对这一部分还知之甚少。
两信号系统的最佳结合是通过“杀伤”和“不杀伤”信号之间的平衡来决定 NK细胞是否摧毁靶细胞。
那么,你是如何认为让NK细胞杀伤不表达MHCⅠ类分子的靶细胞是一个好主意呢?你应该记得,通过检测 MHCⅠ类分子提呈的多肽,CTL能够探测细胞内部是否出了问题?如果一些聪明的病毒关闭了其感染细胞的MHC分子表达,这种病毒感染细胞还会被CTL细胞探测到吗?实际上会的。这种情况下,拥有另一种能杀死表面不表达MHC分子的病毒感染细胞的武器,不是更好吗?
答案是绝对的。这正是 NK 细胞所做的事情,它们可以杀死那些缺乏 MHC分子的细胞。
NK细胞还有另外两个有趣的特征。第一,不同于T细胞需要学会如何如何不攻击自己,NK细胞是一种天才细胞,它们天生就知道该如何做。NK 细胞在看到入侵者时就知道它们是入侵者。值得注意的是,NK细胞更像是综合了杀伤性T细胞和辅助T细胞。类似于CTL一样,NK细胞能破坏被感染的细胞。另外,也类似于Th 细胞,NK细胞也能像发挥细胞因子工厂的作用,实际上,NK细胞就是 IFN-γ的主要供应者之一。
就某种程度而言,NK细胞也类似于巨噬细胞。像巨噬细胞一样,NK细胞也含有包裹破坏性酶和化学物质的颗粒。另外,NK细胞也可能存在几个准备阶段。
静息的NK细胞能产生一些IFN-γ并具有杀伤能力,但是,如果它们被激活,那么它们将会产生更多的IFN-γ,并拥有高效率的杀伤能力。这些杀伤细胞靠什么激活呢?目前已确认一些信号能激活NK细胞,而每一种信号都是在机体遭到攻击时才产生。例如,当NK细胞的表面受体探测到细菌细胞壁的成分入LPS 时,它们就被激活。NK细胞也能被警戒蛋白如 IFN-α/β激活,这些蛋白质是当细胞被特定病毒攻击时细胞释放的。
2.4 先天免疫系统—— 协作效应
为了使先天免疫系统有效地工作,先天免疫系统团队中各成员之间应该进行合作。例如在细菌感染期间,像 LPS 一样的分子结合到 NK 细胞表面的受体上,标志着攻击开始,随后 NK性能通过产生大量的 IFN-γ而作出反应。
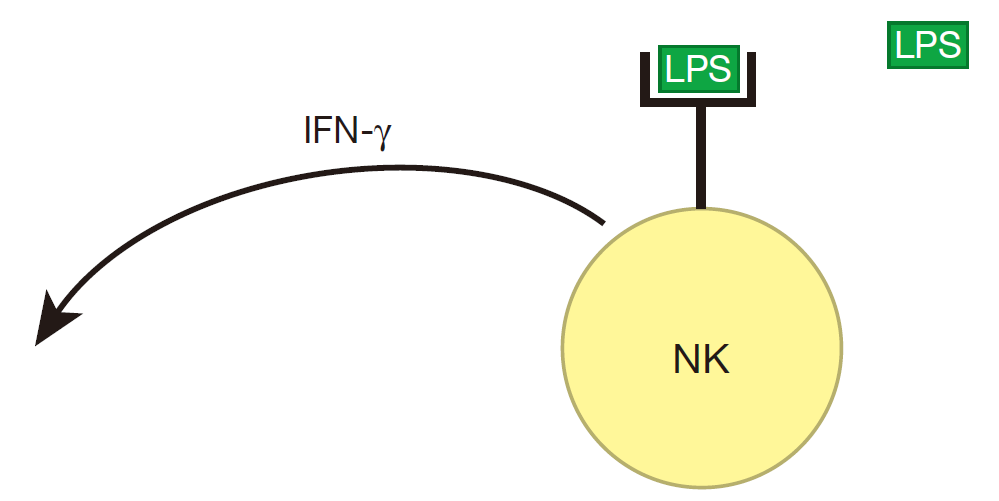
NK 细胞产生的 IFN-γ能致敏巨噬细胞,当巨噬细胞受体也和 LPS 结合时,它们就被超活化。
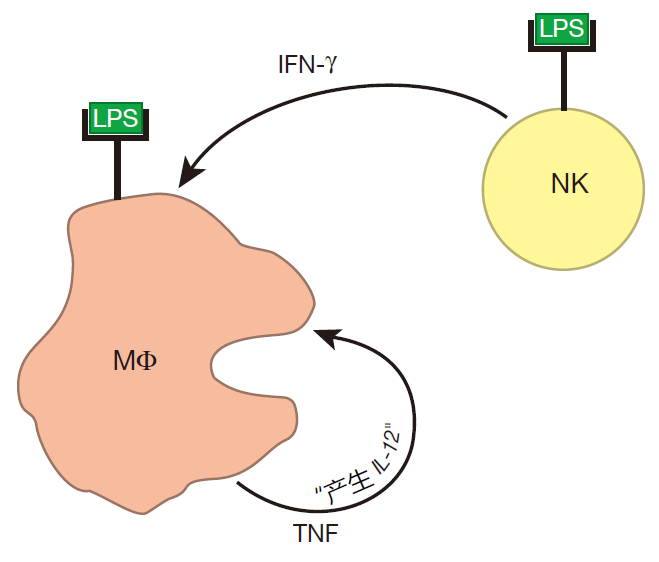
当巨噬细胞被超活化后,就会产生大量的TNF,巨噬细胞表面也有这种细胞因子受体,当TNF和这些受体结合时,巨噬细胞开始分泌 IL-12,TNF 和 IL-12共同影响NK 细胞,增加其 IFN-γ的产量。一旦有更多的 IFN-γ产生,就会有更多的巨噬细胞被致敏。
此外,还需阐明另一些事情。IL-2 是一种由 NK 细胞产生的生长因子,然而,在正常情况下 NK 细胞不表达 IL-2 受体,因此,它们不会因为对这种细胞因子产生应答而增殖,尽管它们是这种细胞因子的制造者。幸运的是,巨噬细胞能够解决这个问题,因为来自于巨噬细胞的 TNF 可以上调 NK 细胞表面的 IL-2 受体表达。因此,NK 细胞能对它们产生 IL-2 作出反应,并开始增殖,增殖的结果导致更多的 NK 细胞迅速参与防御并帮助激活巨噬细胞。
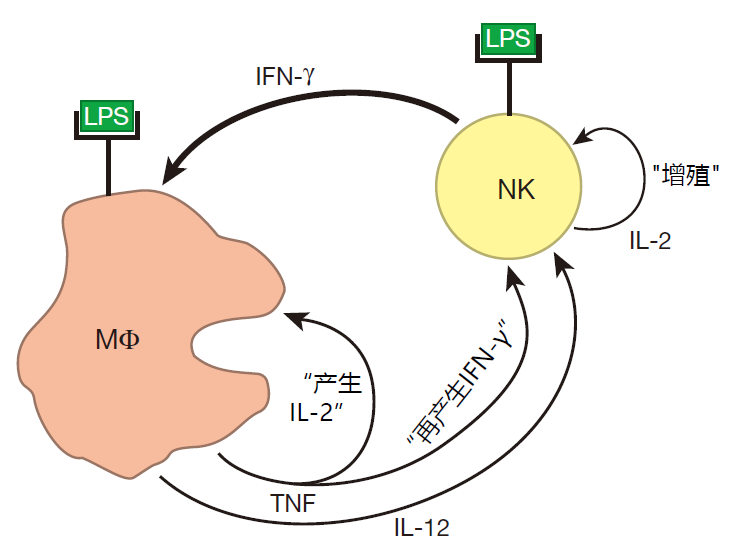
专职吞噬细胞和补体系统也会协同作用。如前所述,补体蛋白片段如 C3b能够标记那些需要被吞噬细胞摄入的入侵者。但在巨噬细胞的激活中,补体调理作用也扮演了重要的角色,因为当修饰入侵者的 C3b 片段和巨噬细胞表面受体结合时,就给巨噬细胞提供了一个类似于 LPS 的激活信号。这真是一个好主意,因为入侵者能被补体调理,但却不会被 LPS 调理。补体系统和吞噬细胞之间的合作不是单向的。激活的巨噬细胞也可以产生几种最重要的补体蛋白:C3,B 和 D。因此,在激烈的战斗中,当补体蛋白被耗尽是,巨噬细胞能为补体系统提供补给。另外,在炎症反应中,巨噬细胞分泌的化学物质增加了血管的渗透性,当这些血管有破损时,更多的补体蛋白被释放到组织中。
吞噬细胞、NK 细胞和补体蛋白之间的相互作用是先天性免疫系统中各成员协同作用的优秀范例,只有通过相互间的合作,先天免疫系统团队中的每个成员才能对入侵者作出快速而强有力的反应。
2.5 先天系统怎样对付病毒
当病毒进入(感染)人体细胞时,它们就接管了细胞结构并用它们来生产大量的子代病毒。最终,这些新制造的病毒从被感染的细胞释放出来,继续感染附近的其他细胞。我们已经讨论了先天系统能用来抵抗胞外病毒的一些武器。例如,补体蛋白能调理病毒,有利于巨噬细胞和中性粒细胞的吞噬作用;通过在病毒表面构建膜攻击复合物,补体蛋白能在包膜病毒(如 HIV-1)上打孔。
尽管先天免疫系统对于抵抗胞外病毒十分有效,但是,一旦病毒进入细胞,先天免疫系统产生和提供的武器就相当有限了。NK 细胞和激活的巨噬细胞分泌像 IFN-γ和 TNF 一样的细胞因子,在某些情况下,这些细胞因子能减少感染细胞产生的病毒量。分泌的 TNF 也能杀死一些病毒感染细胞,特定病毒感染细胞也能直接被 NK 细胞或激活的巨噬细胞杀死。然而,许多病毒一旦进入细胞,它们就能很好地保护自己免受先天免疫系统的武器攻击,这是一个重要的问题,因为,每个病毒感染的细胞都能产生上千个新病毒。
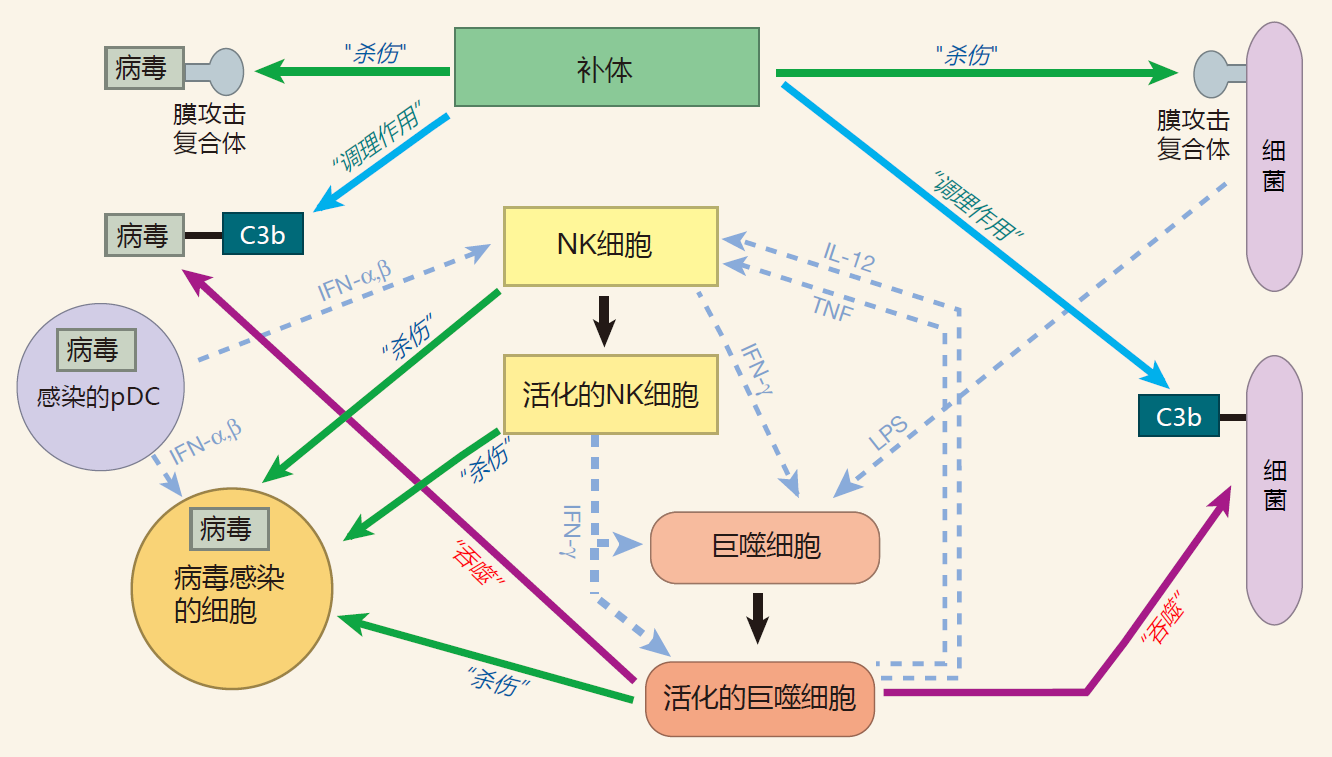
下面的草图显示了补体蛋白、专职吞噬细胞和 NK 细胞能帮助抑制病毒感染,特别是在感染早期。然而,在病毒感染的细胞中,病毒复制很快,而且病毒也是非常聪明的。已经发现了许多病毒逃避先天免疫系统的方法,实际上,机体由此选择了获得性免疫系统来更好地对付这些无赖病毒,这也是我们在后面的第3讲所要讨论的。
2.6 总结图
在这个图中,我总结了本讲中讨论过的一些概念。为了更明晰地表示,我选择巨噬细胞作为专职吞噬细胞的代表,细菌作为人体细胞外入侵者的代表,病毒作为必须进入人体细胞才能完成其生命周期的寄生物代表。在接下来的第3讲中,我将扩展这个图,使之包括来自于获得性免疫系统的成员。