3 B细胞和抗体
3.1 B细胞
在这一讲中,我们将集中讨论获得性免疫系统中最重要的一个成员——B 细胞。像所有血细胞一样,B细胞来源于骨髓,在那里它们从干细胞分化而来。在人的一生中,每天大约产生 10 亿个B细胞,因此即使老年人,也有许多新生成的B细胞。在骨髓B 细胞的早期阶段,它们选择基因片段编码两种组成B细胞受体(BCR)的蛋白质,这些受体继而在B细胞表面占据位置。抗体分子几乎与B细胞受体完全相同,唯一的区别是在其重链末端缺乏锚定于细胞膜的蛋白序列,由于缺乏这种序列,抗体分子被运出 B细胞,自由地到达全身各处,行使自己的职责。我们想谈一点关于选择B细胞受体基因片段的过程,因为我想你将发现它是有趣的,尤其是如果你喜欢博彩游戏的话。
3.1.1 B 细胞受体
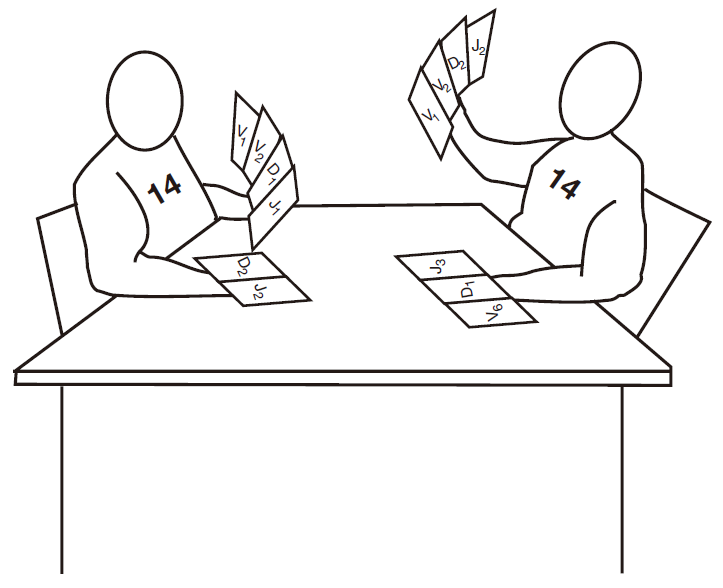
BCR由两种蛋白构成,重链(H)和轻链(L),每种蛋白都是由基因片段组合形成的基因编码。最终形成H基因的基因片段定位于14号染色体上,每个B细胞有两条14号染色体,一条来自父亲,另一条来自母亲,这多少产生了一些问题,因为正如我们以前讨论过,每个B细胞仅产生一种抗体。因为有两套H片段,为了防止B细胞产生两种不同的H蛋白就必须“沉默”1条14号染色体的片段。当然机体可以使1条染色体成为一个虚设物,而让另一条染色体编码H蛋白,但它并没有这样做,这可能很烦人,相反,她则了一个较恰当的方案,我把它图释为两条染色体作为玩家玩牌的游戏,这是一种“胜者至上”的游戏,在这个游戏中,每条染色体都要设法重排基因片段,直到这种排列能够工作。最先完成的玩家将是胜利者。
你回忆一下第一讲中所讲的,通过把四种分隔开的基因片段(V、D、J 和 C)重组在一起,形成编码重链蛋白的基因。这些片段在第14号染色体上线性排列,并有多个拷贝,每种片段不同拷贝仅有轻微的差别。
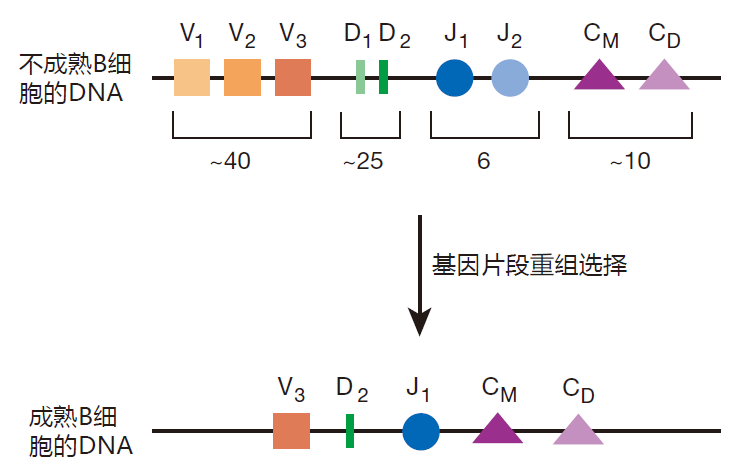
在这种纸牌游戏中的玩家,首先选择 D和J片段中的一个,以缺失它们之间DNA序列的方式将这些片段连接起来,然后再从许多V片段中选择一个,再通过缺失它们之间的DNA,将该片段连接到DJ片段上。紧靠重排后J片段的是一串编码各种恒定区的基因片段(C)。通过缺失,IgM和 IgD的恒定区被用来产生BCR,因为它们位于线性排列的前端。免疫学家把这种基因片段的连接称之为基因重排,但它实际上更多的是切割和粘贴而不是重排。无论如何,结果是选择了V、D、J基因片段和恒定区基因片段最终互相毗邻在染色体上。
接下来,必须对重排基因进行检测。检测什么呢?正如你知道,当核糖体遇到三个终止密码子中的任何一个时,蛋白质翻译将停止,因此如果基因片段恰好正确地连在一起(阅读框正确),蛋白质翻译体系就遇到终止密码子,并终止蛋白质翻译于H中部的某个位置,如果发生这样的情况,就产生一些无用的蛋白质片段。事实上,你可以推算出每个玩家仅有1/9 的机会装配出能产生全长H蛋白的基因片段组合。免疫学家把这种基因片段组合称为“产生性重排”。如果在玩这个游戏时,其中一条染色体最终发生了产生性重排,就成功产生了 H蛋白,并被转运到细胞表面,在该处把“游戏结束”的信号传递给另一条失败的染色体。
有关这样传递信号以及如何终止另一条染色体上基因重排的准确机制仍有待研究。
因为每个玩家 有1/9的成功机会,你或许会想如果两条染色体基因重排后都不能形成产生性重排会怎样,其结果是B细胞死亡,对,它自杀了,这是一个高风险的游戏,因为不能表达受体的B细胞是完全无用的。
如果重链重排是产生性的,轻链“玩家”将进一步进入游戏。游戏规则与重链游戏相似,但必须通过第二次检测,即产生的重链和轻链必须正确地结合在一起才能产生完整的抗体。如果B细胞不能产生性重排生成重链和轻链;或者重链和轻链不能正确配对,B细胞就自杀。因此每个成熟的B细胞产生且仅产生一种BCR或抗体,而这种BCR或抗体仅由一种重链和轻链组成。因为这种用于产生最终重链和轻链基因的混合和匹配策略,使不同的B细胞上的受体如此多样,以至于它们能选择性地识别任何可能存在的有机分子。当你意识到有如此多的有机分子存在时,像这样一个简单的计划可能会创造如此多样的BCR,确实会令你震惊。
3.1.2 BCR如何传递信号
免疫学家把一个给定的BCR所识别的抗原称之为同源抗原,把同源抗原上BCR真正结合的微小区域称之为表位。例如,如果一个B细胞的同源抗原碰巧是流感病毒表面的一个蛋白质,那么表位将是蛋白质与BCR 结合的部分(通常为6-12个氨基酸)。当BCR识别了与它匹配的表位时,它必须把这种识别信号传递到细胞核,并在这里开启或关闭激活B细胞的基因。但是这种BCR如何把发现了它的表位的信号传递到细胞核?初一看,这似乎涉及到许多问题,因为从这张图看,跨过细胞膜进入细胞内部的重链部分在长度上仅为几个氨基酸,太短而不能传递任何重要的信号。
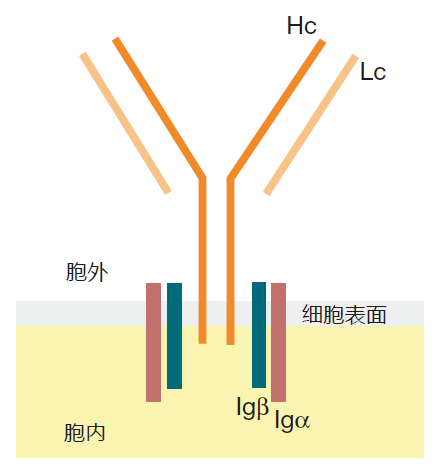
为使BCR细胞外部分能传递它所看到的信号,B细胞上装配了两种辅助蛋白:Igα和 Igβ,它们与重链蛋白相关,并穿过B细胞膜直到细胞内。这样,完整的B细胞受体实际上包括两部分:细胞外识别抗原但不能传递信号的H/L部分以及能传递信号的 Igα和Igβ蛋白,但后者对细胞外发生的一切一概不知。
为了产生信号,B细胞表面上许多 BCR必须聚集在一起。当BCR结合在某一单一抗原的多次重复的表位(如一个蛋白质其一段氨基酸序列重复多次)上时就形成这种BCR簇。
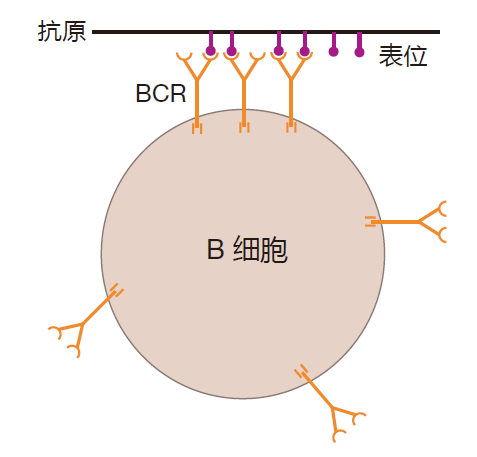
BCR也可能通过结合至入侵者表面聚集在一起的单个抗原上的表位上而被簇集。如大部分细菌和病毒表面含有一些不同蛋白质的许多拷贝。因此,如果BCR识别这些蛋白质中的一种蛋白质表位,那么许多的BCR就可能被簇集在入侵者上。最后,通过结合到簇集在一起的抗原表位(例如一簇蛋白质)上,BCR也可能被聚集在一起。不管怎样完成这个过程,BCR的簇集或交联对B细胞的激活是必须的。下面讲述原因。
信号分子Igα和Igβ的尾部可与细胞内的酶相互作用。当足够强的这种相互作用聚集在一个区域时,就启动把“BCR参与”的信号传递到细胞核的酶链反应。
因此这种传递信息的方式就是把许多的Igα和Igβ分子集中在一起,这就是BCR簇集所做的。BCR簇集使足够多的Igα和Igβ分子聚集在一起,并启动了传递“BCR参与”信号的酶链反应。因此,BCR交联是关键。
除B细胞受体及其相关的信号分子 Igα和 Igβ外,B细胞表面含有另一种蛋白质也可能在信号传递中起着重要的作用。这种蛋白质是一种受体,它能结合正在修饰入侵者的补体片段,结果是,对被调理的抗原而言,B细胞上有两种能与其结合的B细胞受体——识别抗原上特异性表位的 BCR和识别被修饰抗原的补体受体。当这发生时,被调理的抗原就作为一个夹子,在B细胞表面上把BCR和补体系统夹在一起。
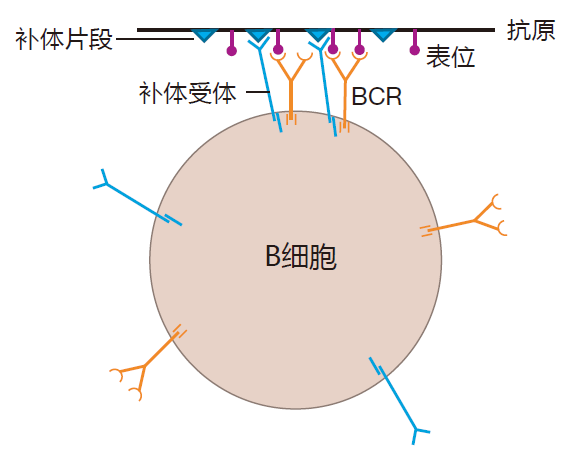
当BCR和补体通过被调理的抗原以这种方式交联时,BCR输送的信号被大大地放大了。在实践中,这意味着把“受体参与”的信号传递到核所必须簇集的BCR数目至少减少至 1/100。因为补体受体对信号传递有如此神奇的影响,它被称为共受体。在进攻的起始阶段,当可与B细胞受体交联的抗原的数量有限时,共受体的功能尤为重要。通过B细胞的共受体识别被调理的入侵者也有助于使 B细胞对先天免疫系统认为是危险的抗原更加敏感。这就是先天免疫系统“指导性”功能的一个很好的例子。确实,一般来说,入侵者是否危险通常是由先天免疫系统决定,而不是由获得性免疫系统决定的。
3.1.3 怎样激活B细胞
为了产生抗体,必须首先激活B细胞。大多数B细胞从来没有遇到它们的同源抗原,这些细胞通常被称之为原初型B细胞。例如B细胞能识别天花病毒,但它可能碰巧存在于从来没有接触天花病毒的人体内。相反,把已经遇到它们的同源抗原的B细胞称之为“有经验的”B细胞。尽管无论原初型的B细胞还是有经验的B细胞一定要被激活,但激活这两种类型的细胞的规则有些不同,因此我们需要分别集中加以讨论,让我们先以原初型的B细胞开始。
激活原初型B细胞需要两种信号。第一种信号是B细胞受体和相关的信号分子的簇集,然而,仅仅交联其受体是不足以激活B细胞的,因此需要第二种信号,免疫学家称之为共刺激信号,通常由辅助性T细胞提供(T细胞依赖的激活)。研究最清楚的共刺激信号是B细胞和Th细胞间直接的相互作用。在激活的Th细胞表面有称之为 CD40L的蛋白,当CD40L插入(连接)B细胞表面上的CD40蛋白时,就开始输送共刺激信号。
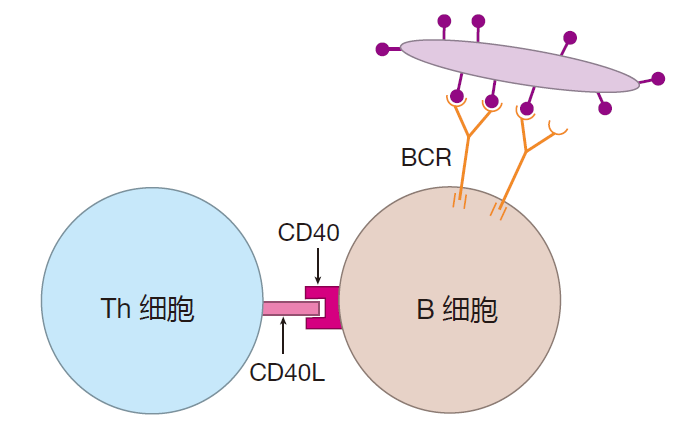
CD40和CD40L之间的相互作用对激活B细胞是十分重要的,如果编码这两种蛋白质的基因中任何一种发生基因缺陷,人类都不能产生依赖于T细胞的抗体应答。
在对特定抗原的应答中,原初型B 细胞也很少或不需要T细胞的帮助就被激活。这些抗原通常具有重复的表位,这些表位能够与许多B细胞受体交联。事实上,大量B细胞受体的簇集可以部分取代CD40L的共刺激作用。在许多细菌的表面发现的糖分子就是这些抗原很好的一个例子。一个糖分子由很多重复单位构成,很像串在一起的珠链。如果把每个珠子看作BCR识别的表位,珠链就可以把许多BCR簇集在一起,并启动B细胞的激活。当然,这种激活是抗原特异性的,只有那些具有能识别重复表位受体的 B细胞将被激活。
还有另一种完全不同的激活B细胞方式,它也是不依赖于T细胞的,在这种情况下,称之为“分裂原”的抗原与B细胞表面的分子而不是细胞受体结合,并聚集这些分子,从事这些分子相关的BCR也簇集在一起,这种“多克隆”的激活是不依赖于被BCR识别的同源抗原的,BCR仅仅是受骗才赶过来的。在这种情况下,单一的分裂原可以激活许多具有不同特异性的许多不同的B细胞。确实,分裂原是免疫学家所喜爱的工具,因为它们能被用于同时激活整个B细胞库,更便于研究激活过程中所发生的事件。
构成某些寄生虫表面的具有高度重复的结构是分裂原很好的例子。在B细胞表面具有能识别这些结构的“分裂原受体”。当寄生虫感染是,B细胞表面的分裂原受体彼此靠近,并集中于寄生虫表面上,当分裂原受体以这种方式聚集时,BCR也被聚集,结果导致在其表面无识别寄生虫BCR的B细胞被多克隆激活。
为什么免疫系统要通过激活那些在表面具有完全不相关受体的B细胞而对分裂原(例如寄生虫表面)进行应答呢?毕竟,那些B细胞对防御寄生虫将是无用的。答案是,这并不是免疫系统设计好要做的多克隆激活完全是非自然的。
通过激活一群产生不相关抗体的B细胞,寄生虫妄图分散免疫系统的注意力,使之不能集中于现有的工作-—破坏寄生虫入侵,因此B细胞的多克隆激活实际上是免疫系统出错的一个例子,这个论题我们将在另一讲中以较长的篇幅讨论。
我在早先提到获得性免疫系统被设计出来,使得必须有两把“钥匙”来激活它,这就像一个保险柜,其中一把钥匙是特异的。对B细胞而言,这把特异性钥匙将是B细胞受体的交联。激活所需的第二把钥匙是非特异性的,即同一把钥匙对所有的B细胞都起作用。在依赖T细胞的激活中,第二把钥匙以共刺激分子CD40L的形式插入B细胞表面的CD40蛋白,通过这“两把钥匙规则”,建立一个保险机制,由一组细胞而不是仅有一个细胞决定是否被激活。
但不依赖于T细胞的B细胞激活又是怎样的呢?它与这“两把钥匙规则”相矛盾吗?确实,许多BCR可能与重复的表位交联,但第二把钥匙在哪里?这丢失的第二把钥匙困扰了免疫学家很长一段时间,因为通过简单地识别仅有重复表位的靶抗原来激活B细胞似乎太危险了,因此直到最近发现不依赖于T细胞的激活确实需要两把钥匙后,免疫学家才感到如释重负。
通过仔细的实验,免疫学家已证实当B细胞受体被交联时,B细胞开始增殖。然而,B细胞增殖后,它并不分泌任何抗体,除非它接收到第二种信号,这第二种信号是什么呢?对不依赖于T细胞的激活来说,第二种信号就如像 IFN-γ的战斗细胞因子,它是继续进攻的明确信号。这就意味着如果B细胞识别了仅有重复表位的分子,例如你自己的DNA,虽然B细胞增殖,但幸运的是并不产生抗DNA的抗体。这是为什么你的免疫系统不参与抵抗你自己DNA的原因——因为没有具有战斗力的细胞因子可以通过共刺激信号。另一方面,如果先天免疫系统要对抗细菌感染,B细胞识别位于入侵细菌表面的、具有重复表位的糖类抗原分子,因为先天性免疫系统产生的战斗信号(如 IFN-γ)提供了B细胞完全激活所需的第二信号,B 细胞将会产生抗体。
因此,在对不依赖T细胞的抗原反应中,B细胞能够直接追随先天免疫系统,不等Th激活就进入免疫阵地。最后的结果是在不依赖T细胞的情况下,对激活B细胞的入侵者迅速产生抗体应答。
但是这里还有更重要的因素,因为T 细胞只识别蛋白抗原,如果所有B细胞的激活都需要T细胞的协助,那整个获得性免疫就将完全集中于蛋白质上。
但事实并非如此,因为大多数入侵者的表面具有在人体细胞表面未发现的糖类和脂类分子,这些独特的糖类和脂类分子组成了被免疫系统识别的很好靶点。因此,通过允许一些抗原不需要T细胞的帮助而激活B细胞,机体就做了一件很精彩的事情——它增加了针对抗原的获得性免疫反应的多样性,不仅包括蛋白质,而且包括糖类和脂类分子。
正如我所提到的,激活原初型B细胞和有经验的B细胞是不同的。事实上,一旦B细胞被激活后,它就记住了那次经历,当它再次识别自己的同源抗原时,再次激活所需要的抗原量就远没有首次激活所需要的量大。最新的观点认为在再次激活时,需要识别同源抗原,但至少在某些情况下,B细胞与Th细胞之间的物理性接触是不必要的了。
为什么首次激活B细胞较难但再次激活就相对容易呢?这样一个系统的存在具有什么优势呢?很明显,激活原初型B细胞的是比较难控制的,因为当真正受到威胁时,你仅仅想激活获得性免疫系统。因此,对于原初型B细胞需要自动保险激活。另一方面,你又期望在记忆细胞库中,找到大量在你第一次暴露于抗原(如天花病毒)后永久存在的有经验的B细胞。这些“合法的”B细胞在首次激活过程中,已经通过严格的“两把钥匙”选择,可以保护再次受到攻击。事实上,正是这些特别的B细胞,当你再次受到攻击时,你期望可以迅速激活它们,因此,使它们容易再次被激活是非常有意义的。
当B细胞被激活时,它们在表面表达新的蛋白质。IL-2受体就是其中之一,IL-2是一种能刺激B细胞增殖的生长因子。这意味着B细胞的激活使它们能够接收启动增殖的细胞因子信号。这种激活与增殖的耦联形成了克隆选择的基础——只有那些已经识别了它的同源抗原并且已经被激活(选择部分) 的B细胞将对生长因子应答,并发生增殖,形成具有完全相同BCR的B细胞克隆。因为Th是生长因子如IL-2的主要提供者,对产生“选择性”的B细胞克隆来说,通常需要T细胞的辅助。
一旦B细胞被激活,并通过增殖后建立起自己的数量群,它们就准备进入自己生命的下一个阶段——成熟。成熟可以粗略地分为以下3个阶段:类别转换,在这个阶段,B细胞可能改变它产生的抗体类型;亲和成熟,在这个阶段,编码B细胞受体的重排基因可能经历突变和选择来增加BCR对其同源抗原的亲和力;职业决定阶段,在该阶段,B细胞决定是称为抗体工厂(浆细胞)还是称为记忆B细胞。这些成熟阶段准确的次序也可以变化,一些B细胞可以跨过一个阶段或一起跨过更多的阶段。
3.1.4 类型转换
B细胞在骨髓中产生后,它们重排编码重链和轻链蛋白的基因,在其表面展示两种类型的抗体——IgM 和 IgD。这是年轻B细胞的BCR,通常称之为sIgM和sIgD,这里“s”代表“表面”。有趣的是,产生sIgM和 sIgD重链的mRNA是相同的,只是剪接方式不同,即一种剪接方式产生出M型的恒定区,而另一种剪接方式则产生D型的恒定区。在人体循环系统中,IgD抗体仅仅占抗体的很少的部分,还不清楚这些抗体是否具有显著的功能。相反,存在于原初型B细胞表面的sIgD对B细胞激活是重要的,实际上,平均每个原初型B细胞表面的IgD都较IgM多。
当B细胞离开骨髓时,因为还没有被激活,它不分泌抗体。这种原初型B细胞必须首先寻找它的同源抗原。如果B细胞找到同源抗原,并且接受到其所需要的共刺激信号,B细胞将被激活,一旦被激活,B细胞就准备产生默认的抗体类型——IgM。然而B细胞也有机会将其产生的抗体类型由 IgM 转变为 IgG、IgA、IgE中的任何一种。你应该还记得抗体的类型是由重链的恒定区(Fc片段)决定的,即抗体分子的“尾部”。位于编码 IgM恒定区的基因片段旁边的是编码IgE、IgG、IgA的恒定区基因片段。因此,所有B细胞要进行类别转换就必须切掉IgM的恒定区,并与其他任何一种抗体的恒定区相连(即缺失它们之间的DNA)。在染色体上,位于恒定区基因片段之间的是特定的类型转换信号,它允许在该处发生切割和粘贴。例如,下面所示的是B细胞由IgM的恒定区(CM)转换为IgG的恒定区(CG)时所发生的情况。
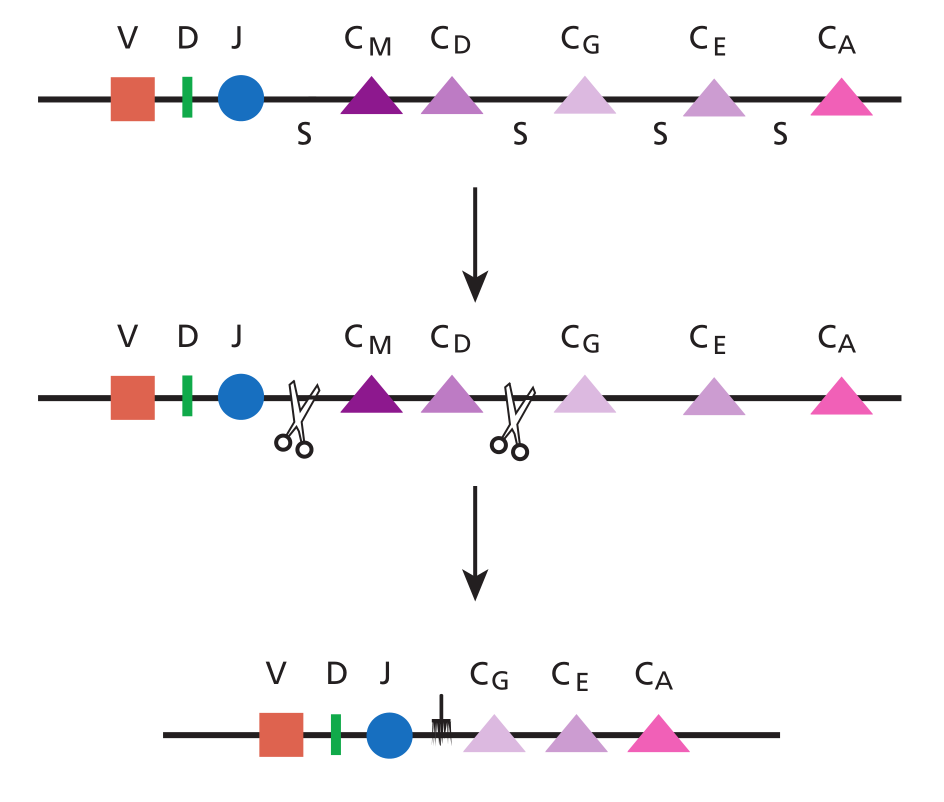
类型转换的最终结果是尽管抗体与抗原的结合部分仍然是相同的(Fab),但抗体的尾部变了。因为决定抗体功能的是其恒定区,所以这是一个重要的改变。
3.2 抗体及其功能
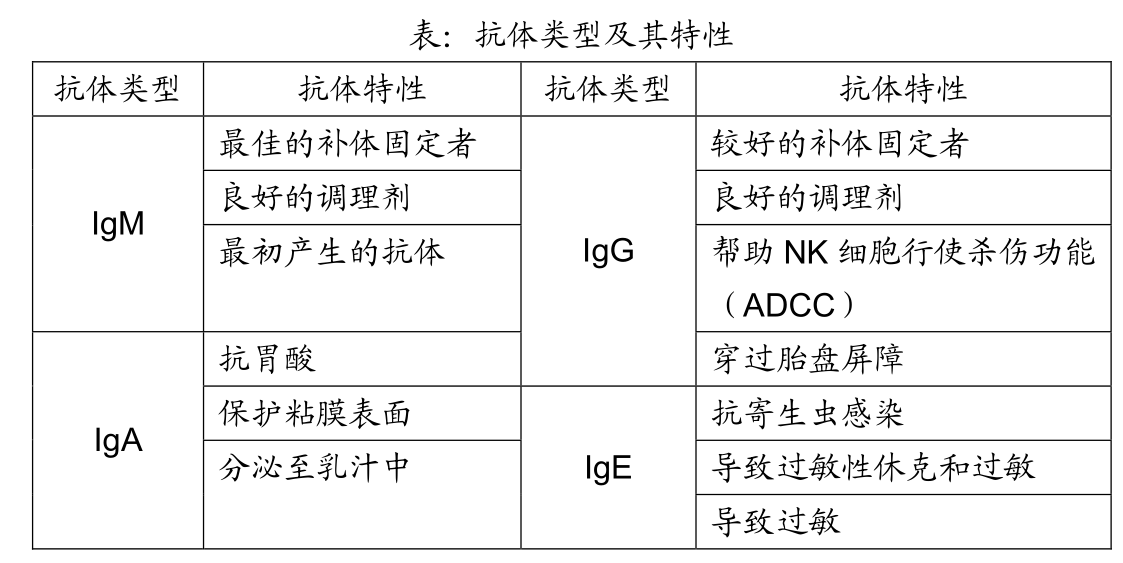
让我们看一下4种主要类型的抗体——IgM、IgG、IgA、IgE。正如你将看到,由于它们各自的恒定区(Fc)具有独特的结构,所以每种类型的抗体,都有其特定的职责。
3.2.1 IgM抗体
IgM型抗体是最早形成的抗体类型,甚至“较低等”的脊椎动物(我向动物权益保护者道歉)也具有产生IgM抗体的获得性免疫系统。因此,当人体的原初型B细胞首先被激活时,它们主要产生IgM型抗体,这是合理的。你可能还记得第一讲中所描述的 IgG抗体看起来像什么。
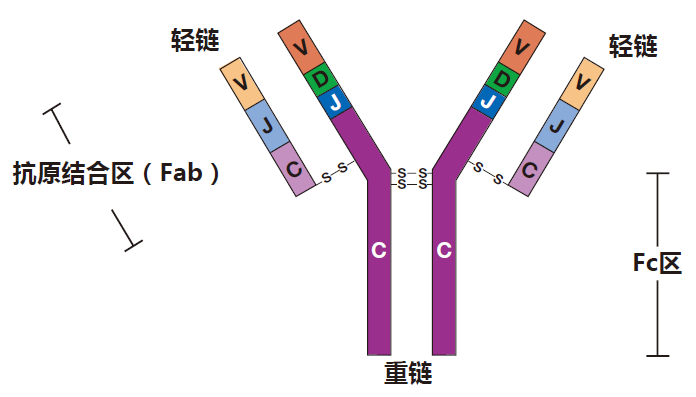
一个IgM抗体就像是把5个IgG分子粘在一起的,它确实是巨大的。
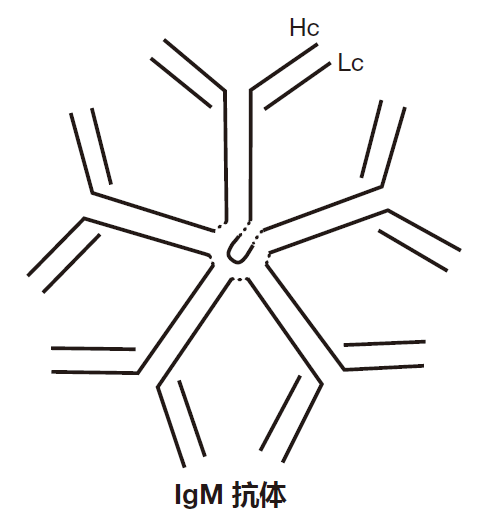
因为IgM抗体对于激活补体级联反应是非常有用的(免疫学家称之为“固定”补体),所以在感染的早期产生如此巨大的IgM型抗体是相当明智的。这里讲述其如何作用。
在血液和组织中,一些补体蛋白(约30种)聚集在一起形成一种称之为 C1的大分子复合物。尽管很大,但这种蛋白复合物还不能激活补体级联效应,因为它结合到抑制剂分子上。然而,如果2个或更多的C1复合物聚集在一起,它们的抑制剂就脱离,然后C1分子才能启动产生C3转换酶的级联反应。这时补体系统就开始工作,你应该还记得上一讲中所提到的内容,C3转换酶把C3转换为C3b,建立了产生越来越多C3b的扩大环。因此,通过这种经典途径(抗体依赖的)激活补体的方式能够将两个或更多的C1分子聚集在一起,这正是 IgM抗体所能做的。
当IgM抗体的抗原结合区结合到入侵者时,C1复合物就能结合到抗体的Fc段。因为每个IgM抗体有5个聚在一起的Fc区(这一点很重要),两个C1复合物能够结合同一个IgM 抗体的Fc区,使复合物聚集到足够近,从而启动了补体级联反应。因此,这一事件的顺序是:IgM抗体结合到入侵者上,一些C1分子结合到IgM 抗体的Fc区,这些C1分子就在入侵者表面启动补体链式反应。
这是先天免疫系统(补体蛋白)与获得性免疫系统(IgM抗体)共同破坏入侵者的一个很好的例子。事实上,当免疫学家第一次发现,如果抗体被其他蛋白(补体蛋白)补充抗体就能更有效地破坏入侵者,他们就创造了“补体”这个术语。
在上一讲种我们所讨论的补体激活的旁路(自发)途径完全是非特异性的——任何未受保护的表面都公平地进入“游戏”。相比之下,经典或抗体依赖的途径是相当特异的——只有那些被抗体结合的抗原将被补体靶向攻击。在这种系统种,抗体区分入侵者,补体蛋白做清除工作。
特定的IgG抗体亚型也能够“固定”补体,因为C1能够结合这些抗体的Fc区。然而IgG抗体实际上是无用的,因为每个IgG分子只有一个Fc区,因此要使两个C1复合物足够靠近以启动后面的反应,就需要两个IgG分子在入侵病原体表面紧密结合,这种情况只有当周围有许多IgG时才可能发生。因此在感染的早期,当抗体刚开始产生时,IgM抗体具有比IgG 抗体更大的优势,因为他们能有效地固定补体分子,另外,IgM型抗体通过结合病毒并防止它们感染细胞,而很好地中和病毒,因为这些特性,IgM是预防病毒或细菌感染最好的“第一抗体”。
3.2.2 IgG型抗体
IgG型抗体包括许多不同的亚型,这些亚型的Fc区有轻微的差别,因此具有不同的功能。例如,IgG型抗体的一种亚型IgG3,可以较其他任何亚型更好地固定补体。相似地,IgG1亚型擅长于结合入侵者并调理它们以利于专职吞噬细胞的吞噬,这是因为在巨噬细胞和中性粒细胞表面有能与已结合入侵者的IgG1抗体Fc区结合的受体。
自然杀伤细胞表面有能与IgG3抗体 Fc区结合的受体,这样,IgG3就能够在NK细胞及其结合点的靶细胞之间搭桥,它用自己的Fab区与靶细胞结合,用Fc区与NK细胞结合,这不仅使NK细胞与它的靶细胞靠近,而且其Fc区受体与NK细胞结合后,可以更有效地刺激后者的杀伤活性,这个过程被称之为“抗体依赖的细胞毒作用(ADCC)”,在ADCC过程中,NK细胞杀伤靶细胞,抗体辨别靶细胞。
类似IgM型抗体,IgG抗体也能很好地中和病毒。然而,IgG抗体的独特之处在于它们能够通过胎盘屏障从母体血液进入胎儿血液,这就为胎儿提供了IgG抗体,直到他们出生后几个月能自己产生抗体。由于IgG抗体是寿命最长的抗体类型,半衰期大约为三周,所以能够提供一种长期的保护作用,而IgM抗体的半衰期大约只有一天。
IgG抗体有时也称之为“γ-球蛋白”,如果你有可能暴露于感染原,例如甲型肝炎病毒,医生就可能推荐你注射γ-球蛋白,这些γ-球蛋白的制备是通过收集大量人的抗体,其中至少有一些已感染过甲型肝炎病毒,因而具有了抗病毒抗体,通过这种“借来”的抗体就能中和大多数你接触过的病毒,直到你自己的免疫系统被激活。
3.2.3 IgA型抗体
这里给你提个问题:在人体内最丰富的抗体类型是什么?不是IgG,而是IgA。这个问题问得很有技巧,因为我以前告诉过你,IgG的的确确是血液中最多的抗体。但最终我们体内合成的IgA比所有其他抗体加起来还多。为什么有这么多的IgA?因为IgA是机体保护其黏膜表面的主要抗体,一个人大约有400平方米的黏膜表面需要保护,这些黏膜表面包括消化道、呼吸道、生殖道黏膜,因此,尽管在血液循环系统中没有很多的IgA,但却有大量的IgA在保护黏膜表面,实际上,大约有 80%位于这些黏膜表面下的B细胞会产生IgA抗体。
IgA如此擅长于防御能穿透黏膜屏障的入侵者,原因之一在于每个IgA分子就像被一把“钳”抓在一起的两个 IgG分子。
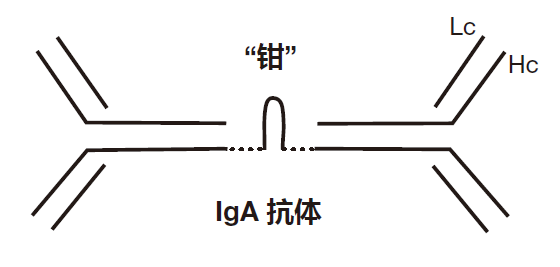
IgA抗体的“钳”型尾赋予了这型抗体一些重要的特性,由于“钳”把它们夹在一起,IgA能通过血液穿过肠壁转运到肠腔,一旦进入肠腔,IgA抗体能够“包裹”入侵的病原体,阻止它们吸附在它们本来能够感染的肠细胞上,另外,虽然每个IgG 分子有两个抗体结合区,但二聚化的IgA分子有四个手结合抗原。因为它们是二聚化的,所以IgA抗体非常有利于把病原体聚集成更大的簇,从而以粘液的形式排出体外。实际上,在粪便中,细菌占了被排泄物的 30%。
IgA抗体独特的尾部结构能抵抗消化道的酸和酶,总之,这些特性使得 IgA能很好地保护黏膜表面。事实上,分泌到哺乳母亲乳汁中的抗体也是IgA型抗体,这些IgA抗体覆盖在婴儿的肠黏膜表面,防御婴儿摄入的病原体。这非常有意义,因为你知道,婴儿喜欢用口接触所有东西,所以他们会从口摄入大量的微生物。
尽管IgA抗体对抗黏膜入侵者非常有效,但它们对固定补体却完全无用,因为C1不能结合到IgA抗体的 Fc 区。由于抗体类型和功能是由其恒定区决定的,缺乏补体固定活性实际上是一件很好的事情,如果IgA抗体也能启动补体反应,我们的黏膜表面将永远处于对付那些侵袭我们黏膜表面的病原体和非病原体的炎症状态,而慢性发炎的小肠却不会出现这样的情况。因此IgA抗体的主要功能是作为被动抗体,阻止入侵者吸附到位于黏膜表面的细胞上,并将这些不受欢迎的客人请出体外。
3.2.4 IgE型抗体
IgE抗体亚型的发现非常有趣。在20世纪早期,一个名叫Charles Richet法国医生与摩纳哥王子 Albert 一同出海,王子向 Richet 提到了一个奇怪的现象,一些人被僧帽水母蛰了后,对毒素产生非常强烈的反应,他建议这种现象或许值得研究。
Richet接受了建议,当他回到巴黎后,他决定做的第一个实验就是产量多大剂量的毒素可以杀死一条狗。不要问我为什么他决定在实验中使用狗,或许在他周围有许多无家可归的狗,或许仅仅是因为他不喜欢小鼠,无论如何,他的实验成功了,他确定了毒素的致死剂量。然而,许多狗因为在第一次实验中没有给予致死剂量的毒素而活了下来。这并不是浪费一只好狗,Richet决定用毒素再次注射这些幸存者,并观察将会发生什么?他猜想这些动物或许对毒素产生了免疫,因为第一次注射可能对第二次注射提供了保护(预防)。你能够想象当所有的狗都死了的时候他是多么的惊奇,即便是那些第二次注射仅用了微量毒素的动物也未能幸免。因为第一次注射产生了与预防相反的作用,Richet用“过敏反应”这个单词来描述这种现象 (ana的意思是相反)。此后,Richet继续研究过敏性休克,并在 1913年因此而获得诺贝尔奖。这里我想我们应该学到的是,如果一位王子建议你研究什么,你或许应该慎重采纳他的意见。
免疫学家现在知道过敏性休克是由肥大细胞脱颗粒所引起的。类似巨噬细胞,肥大细胞是一种位于所有暴露表面(如表皮或黏膜屏障)下的白细胞。就像血细胞一样,肥大细胞的寿命很长,在我们的组织里,它们能够存活数年,等待防御入侵者。近年来的研究表明,在先天免疫防御细菌的过程中,肥大细胞发挥了重要的作用,它们能够吞噬被调理的细菌,也能释放细胞因子募集中性粒细胞和其他免疫细胞到细菌感染的部位,然而,肥大细胞最重要的功能是防御能穿过保护屏障的寄生虫感染。肥大细胞里大量的颗粒包含有各种具有药理活性的化学物质,其中最著名的就是组胺。当肥大细胞遇到寄生虫时,它释放这些颗粒内的物质来杀死寄生虫,然而,出了杀死寄生虫外,肥大细胞脱颗粒也能导致过敏反应,在极端情况下,发生过敏性休克。下面讲述这一切是如何发生的。
把能引起变态反应的抗原称之为过敏原(如僧帽水母的毒素)。当某些人第一次接触过敏原是,因为一些现在还不清楚的原因,产生大量的针对这些过敏原的IgE抗体。肥大细胞表明具有能结合这些IgE抗体Fc区的受体,当发生这种情况时,肥大细胞就像手榴弹一样等待爆炸。
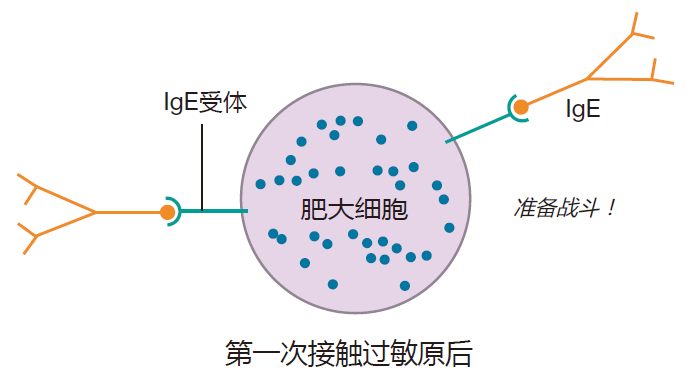
当再次接触过敏原时,已结合于肥大细胞表明的IgE抗体能结合抗原,因为过敏原通常是具有重复序列的小蛋白质,因此过敏原就能交联许多肥大细胞的IgE分子,从而把Fc区受体拖到一起。这种受体的簇集与B细胞受体的交联相似,这些受体的簇集导致了信号传递。然而,这里的信号是脱颗粒,然而肥大细胞将其颗粒释放到你的组织中。
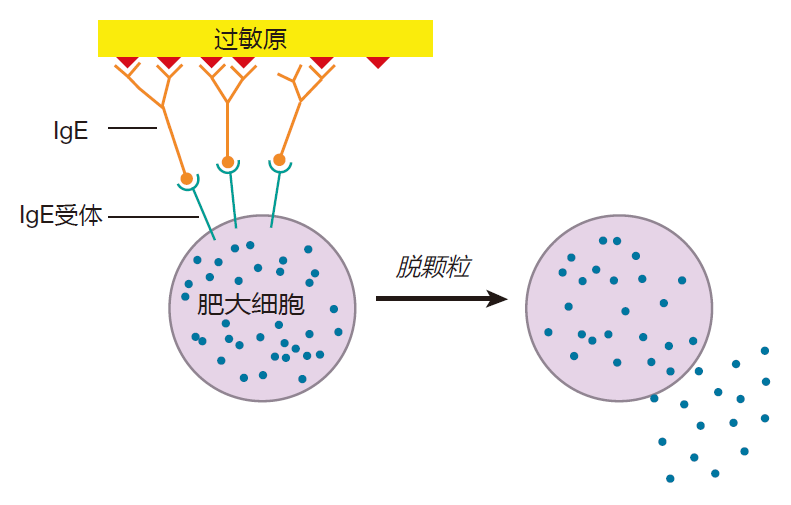
从肥大细胞颗粒中释放的组胺和其他化学物质可以增加毛细血管的通透性,因此液体就从毛细血管流入组织中,这就是为什么当你发生变态反应时,流鼻涕和眼泪汪汪的原因,通常这是局部反应,但是如果毒素扩散到全身,并启动大量的肥大细胞脱颗粒,那情况就变得非常严重。在这种情况下,液体从血管进入组织可能减少血容量,以致心脏不能再有效地泵血,导致心梗。另外,从颗粒中释放的组胺能够导致气管周围的平滑肌收缩,从而导致呼吸困难,严重的可能导致窒息。在这里,我们不必过多担心僧帽水母,我们应该关注的是蜜蜂,因为蜜蜂蛰针中的毒素可能导致某些人致死性过敏性休克,因为这些人针对蜜蜂毒素分泌了大量的 IgE抗体。
这带给我们一个有趣的问题:为什么B细胞允许以任何方式转换它们产生的抗体类型,与古老的IgM的抗体结合不是更安全吗?
假设病毒感染了你的呼吸道,你是否还只想产生一种IgM抗体?当然不,你需要分泌许多的IgA抗体到呼吸道的黏膜中结合病毒,并将它们从你体内清除。另一方面,如果你感染了就是场区(如某种蠕虫),你需要产生 IgE抗体,因为IgE抗体可以导致类似肥大细胞一类的细胞脱颗粒,从而使蠕虫发生生命危险。因此,这个系统的完美在于不同亚型的抗体在防御不同入侵者中有独特的作用。
当你患感冒时,机体使你的免疫系统产生IgA抗体,当你有寄生虫感染时产生IgE抗体,那不是更好吗?是的,事实证明确实是这样的,这里谈一下机体的细节。
当发生类型转换时,由B细胞所遇到的细胞因子来调控抗体类型的转换:特定的细胞因子或几种细胞因子组合影响了B细胞转换为一种或另一种类型。
例如,如果B细胞发生类型转换时周围环境富含IL-4、IL-5,它们优先从 IgM向IgE转换,这正好可以预防寄生虫感染;另一方面,如果周围有许多IFN-γ,B细胞就发生类型转换产生IgG3型抗体,这对于细菌和病毒感染非常有效;又或者,如果在类型转换时存在TGF-β,B细胞优先从 IgM转向产生IgA,这可以对抗普通感冒。因此为了确保抗体反应与入侵者相匹配,当B细胞发生类型转换时,机体必须使其周围产生正确的细胞因子。但她是如何完成的呢?
你可能还记得,辅助T细胞是指导免疫反应的指挥细胞,它们发出指令的方法之一是产生细胞因子影响B细胞产生正好能防御入侵者的抗体类型。为了弄清Th细胞这样知道该产生哪一类型的细胞因子,在接下来的二讲中,我们讲讨论抗原提呈和T细胞激活。但现在我只能给你一个底线:为了对Th细胞产生的细胞因子作出应答,B细胞必需将其产生的IgM抗体向其他类型的抗体转换,这是的获得性免疫系统有可能对每种病原体产生特定的抗体应答,不管这些入侵者是细菌、流感病毒还是寄生虫,还有什么比这样更好呢?
3.3 体细胞高突变
如果类型转换还不足够,那么在B细胞成熟过程中还会发生另一件十分有趣的事件。通常,在人体细胞中,DNA整体突变率是相当低的,在每个DNA复制周期中,每1亿个碱基中大约仅有一个会发生突变。突变率必须这样低,否则我们最终看起来像星球大战中的人物——具有三只眼睛和六只耳朵。然而,在B细胞染色体非常有限的区域(包含 V、D、J基因片段的区域)里会发生相当高的突变率。事实上,在这一区域,已测到在每一代中,大约每一千个碱基中就有一个发生突变,我们称这里发生的是非常严重的突变!这种高突变率叫做“体细胞高突变”,发生与 V、D、J基因片段选择后,通常在类型转换后发生。因此,在B细胞成熟过程中,体细胞高突变是发生相对晚的事件,并且持续产生IgM型抗体的B细胞通常不经历体细胞高突变。
体细胞高突变改变了部分的重排抗体基因,该部分基因编码抗体的抗原结合区,根据突变的程度,有3种可能的结果:抗体分子与其同源抗原的亲和力或许保持不变,或者增加,或者降低。有效的结果是为了使成熟的B细胞可以继续增殖,它们必须结合于其同源抗原而不断被重新刺激。因此,由于那些BCR突变后具有更高亲和力的B细胞更易受到刺激(因为BCR结合的更好),它们比具有低亲和力BCR的B细胞更易增殖。也正因如此,体细胞高突变的结果是产生了更多的B细胞,这些B细胞的BCR对其同源抗原具有高亲和力。
通过体细胞高突变,改变BCR的抗原结合区:通过结合与增殖筛选那些增加了BCR与抗原结合能力的突变体,BCR可能被“优化”。这样的结果是选择出了具有对同源抗原具有较高亲和力受体的B细胞,这个过程称之为亲和力突变。
因此,B细胞通过类型转换可能改变了他的恒定区,通过体细胞高突变改变它们的抗原结合区,这两种修饰的结果是产生了能更好对付入侵者的B细胞,但是不需T细胞帮助激活的B细胞通常不经历体细胞高突变和类型转换。
3.4 B细胞面临的职业选择
B细胞成熟的最后一步是职业选择。这不可能太复杂,因为B细胞实际上仅需在变成浆细胞或记忆细胞两条途径之间作出选择。浆细胞是抗体工厂,如果B细胞决定变成浆细胞,它通常要到达脾脏或回到骨髓中去,产生分泌型的BCR——抗体分子。浆细胞每秒钟大约可以产生两千个抗体分子,一个浆细胞产生如此多的抗体,使免疫系统能够防御像细菌或病毒这些增殖十分迅速的病原微生物入侵。然而,这种扩大的效应的结果是,大部分B细胞仅能存活几天。
尽管B细胞的另一个职业选择——变成记忆B细胞——或许并不像决定变成浆细胞那样惹人注目,但它却是极其重要的,记忆B细胞记住了第一次入侵的抗原,可以防御以后的攻击,记忆B细胞通常经历类型转换,产生专门抵御它们记忆中的外来入侵者的抗体类型;另外,大多数记忆B细胞经历了体细胞高突变,因此它们具有高亲和力的 BCR,可以对感染初期的少量抗原产生应答;最后,记忆B细胞比原初型B细胞具有低的激活需求,因为这些特性,记忆B细胞随时准备去抵御第二次进攻。
记忆B细胞能够对感染产生长期的免疫。例如,1781 年瑞士商人将麻疹病毒带到了与世隔绝的法罗岛,而在 1846年,当另一只载有感染了麻疹病毒的海员到达该岛时,那些大于 64岁的人没有发病,这是因为在它们的体内还有麻疹病毒的抗体。持续如此长时间的B细胞免疫是怎样维持的,这是当前一直争论的。即便是寿命最长的抗体(IgG亚型)也只有大约3周的半衰期,因此必须连续产生抗体以提供长期的保护。有一些证据显示,记忆B细胞能够存活较长的时间,另一些实验则表明,记忆B细胞的寿命相对较短,但当它们被抗原重新刺激时,B细胞连续组织,这可能是攻击后残留的抗原或与入侵者相似的另一种抗原重新刺激了记忆B细胞,按照这种说法,这正是那些能够产生具有免疫能力的抗体的原初记忆B细胞的子代。
免疫学家还不能描绘出B细胞怎样选择变成记忆B细胞或浆细胞,然而,他们知道,辅助T细胞表面的CD40L 与B细胞表面的CD40之间的相互作用对于记忆细胞的组织具有对于类型转换和体细胞高突变一样的重要性。这种对CD40L与CD40相互作用的需要有助于解释为什么当在无 T细胞帮助时,B细胞激活,但却不能产生记忆B细胞以及为什么不依赖于T细胞的激活通常产生IgM抗体,而无需被体细胞高突变所优化。
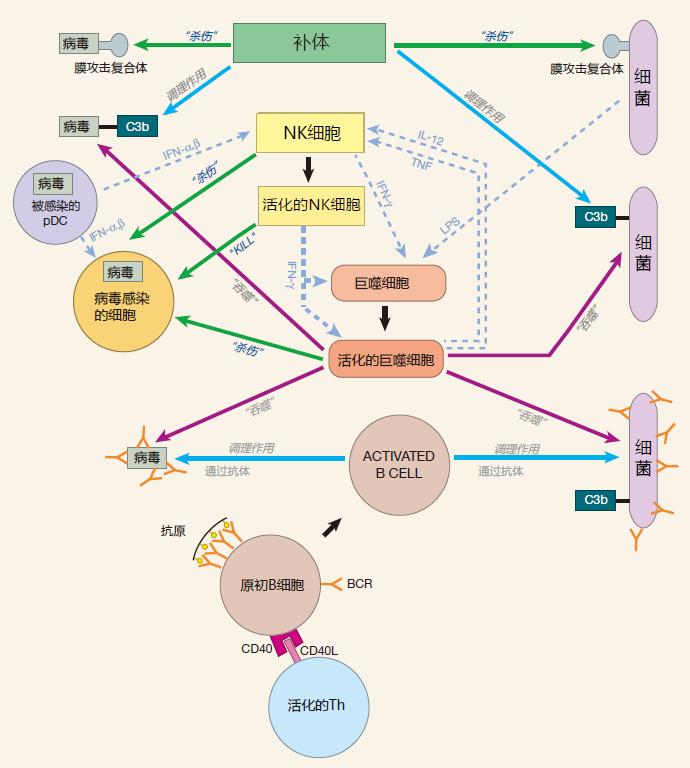
3.5 总结图
这张图包括了上一讲中的先天免疫系统以及我们在这里所讨论的B细胞。