
3.1 长寿和衰老进化理论的基础
长寿和衰老的进化理论植根于进化生物学的基本概念之中。在本节中,我们将简要探讨对长寿和衰老相关进化模型的发展产生重大影响的一般进化主题。
魏斯曼在体细胞和生殖细胞之间建立了分离
1882年达尔文去世时,生物科学界尚未完全接受自然选择进化。尽管当时大多数生物学家都认为达尔文毫无疑问是正确的,但该理论仍然存在许多漏洞,无法被完全接受。其中一个问题是性状在世代之间传递的机制。达尔文时代的大多数科学家认为,体细胞将遗传特性直接传递给生殖(性)细胞或配子。直到伟大的德国理论家奥古斯特·魏斯曼(August Weismann,1834-1914)证明了这一点,体细胞和生殖细胞分裂的概念才得以确立。这为衰老理论奠定了基础。在一项实验中,魏斯曼交配了22代在生殖年龄之前尾巴被剪掉的老鼠。每对交配的老鼠都有尾巴(图3.1)。这项实验表明,体细胞与性细胞没有通讯,只有配子才能将遗传的化学物质传递给下一代。
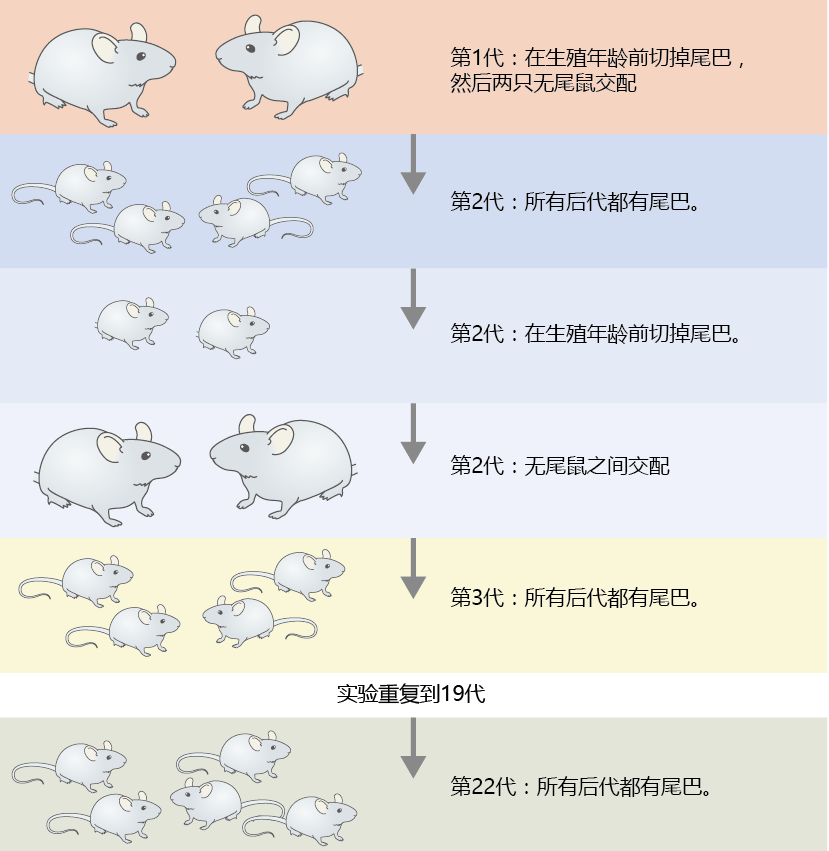
图3.1 魏斯曼的实验建立了生殖系(性细胞)和体细胞(体细胞)之间的分离。魏斯曼发现,在繁殖开始前去掉老鼠的尾巴对后代一代又一代的尾巴外观或长度没有影响。
体细胞和生殖细胞之间的分离向魏斯曼暗示了生物体内的分工:体细胞的存在仅仅是为了支持生殖细胞及其传递遗传“物质”的功能。基于这一证据,魏斯曼从理论上推断,躯体的工作是确保个体的寿命足以繁衍后代。一旦完成这项工作,就不再需要胞体了,生物体就会衰老和死亡。正如本章后面所讨论的,魏斯曼关于衰老的思想构成了后来被称为权衡(trade-off)假说的基础,该假说认为成功繁殖的成本是必死性。
魏斯曼提出,衰老是一种非适应性特征
在韦斯曼关于进化和衰老的第一篇文章中,他指出
……在调节生命的持续时间时,对物种而不是对个体的优势是唯一重要的。这一点对于任何曾经彻底思考过自然选择过程的人来说都是显而易见的。个体寿命的长短对物种来说并不重要,但重要的是个体应该能够为维持物种而努力。这项工作是繁殖或形成足够数量的新个体,以补偿物种的死亡。一旦个体在这项补偿工作中完成了自己的任务,它就不再对物种有任何价值,它已经履行了自己的职责,它可能会死亡。(韦斯曼,1891)
魏斯曼的这篇早期文章表明,衰老是通过自然选择产生的,是一种积极的适应,对物种有利,这是一个被称为群体选择的一般概念。也就是说,选择发生在群体层面,而不是个人层面。魏斯曼最初认为,选择衰老是为了摆脱那些年老无用的个体,这些个体不再具有繁殖能力,但是它们却继续使用有限而宝贵的资源,如食物和水。除去那些对物种的延续没有贡献的无繁殖能力的成年成员,可以让资源优先分配给该群体中繁殖活跃的成员,因此剩余群体的适应度也会提高。
魏斯曼的群体逻辑有助于消除年长、无生育能力的成年成员的物种,这一逻辑具有明显的吸引力,但是此逻辑仍然是一个常见的误解。分子进化研究的证据已经确凿地证明,个体,或者更准确地说,基因是适应的焦点,这正是达尔文最初所相信的。只有生殖活跃的个体才能影响基因组的组成,而这种组成完全是由个体达到生殖年龄和遗传基因的需要驱动的。换句话说,这种基因并不“知道”在以后的岁月里,这个群体需要摆脱老年人。事实上,现在有一些证据表明,更年期(生殖衰老)和延长生殖后寿命可能是一种可以增强体质的适应(框注3.1)。
| 框注3.1 外婆假说与女性寿命 |
|
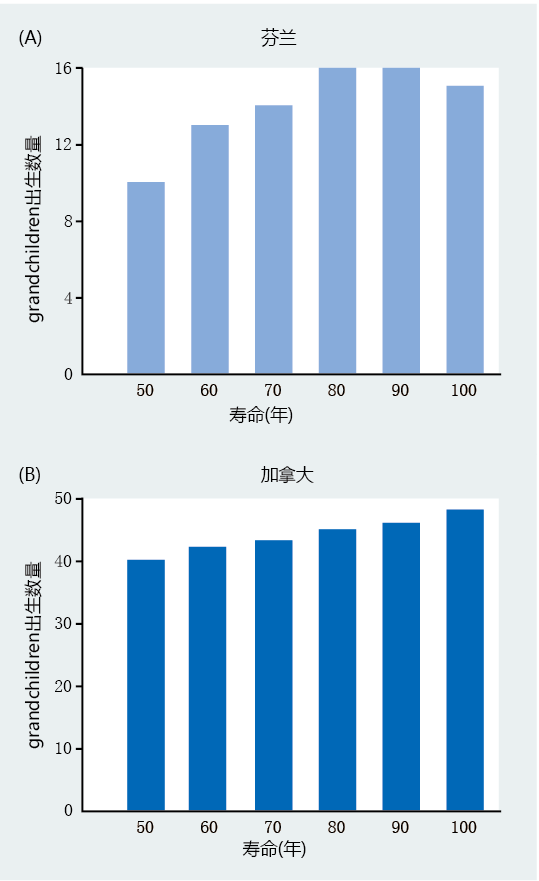
纵观历史,人类外婆一直被描绘成一个大家庭中善良而慈爱的成员,为他们的孙辈提供照顾。许多文化人类学家认为,外孙关系将我们的物种与所有其他物种区分开来,包括非人灵长类动物。一些人类学家现在认为,孙辈和外婆之间的关系超越了简单的情感联系,可能在长寿的进化发展中起到了一定作用。外婆假说假定,在早期社会,外婆对孩子的照顾让他们的女儿生更多的孩子,从而提高了物种的整体适应性。外婆活得越久,女儿生的孩子就越多,身体也就越健康。也就是说,至少在人类身上观察到的较长的生殖后寿命,是因为它有利于生殖成功。 外婆假说是G.C.威廉姆斯理论的延伸,该理论认为更年期可能是一种适应,可以提高物种的整体适应性。威廉姆斯认为,在进化史上,年长的母亲在分娩期间死亡的风险更大,因此无法养活幸存的孩子。这些后代的死亡将导致该物种的适应性下降。然而,如果老年妇女由于更年期而在生理上无法生育,老年母亲可以专注于为已经存在的后代提供资源。类似地,外婆假说表明,由于外婆帮助养育女儿的年长子女,生育后的寿命很长。这将使母亲能够将更多的资源集中在仍然完全依赖她的年轻后代身上。外婆对孙辈的生存至关重要。因此,较长的生殖后寿命可以提高健康水平。 评估18世纪和19世纪收集的人口数据的研究普遍支持外婆假说。研究人员选择了几种健康指标,包括(1)生育更多子女的母亲,(2)生育更早、更频繁的母亲,以及(3)生育间隔更短的母亲。研究人员发现,孙辈的数量与外婆的寿命直接相关(图3.2);外婆的年龄越大,孙子孙女就越多。调查人员还发现,与不包括外婆的家庭相比,当外婆与女儿住在同一所房子里并照顾外孙外孙女时,他们的健康状况得到了改善。基于这些发现,作者们得出结论:“我们的结果有力地支持了延长女性生殖后寿命是适应性的假设,我们的知识首次揭示了女性在超过生殖年龄后生活所带来的实质性健康益处。” 图3.2 女性寿命与下一代孙辈总数之间的关系。数据显示(A)芬兰(1702-1823)和(B)加拿大(1850-1879)。每个人口斜率的增加相当于生育后妇女在50岁以后每10年多生两个孙子。(摘自Lahdenpera M et al.2004.《自然》428:178–181.获得《自然》出版集团的许可。) 其他研究没有发现外婆对物种适应性的“益处”,但已经证明,外婆作为照顾女儿孩子的人缩短了生育间隔。如果一位外婆能够照顾最近出生的孩子,那么母亲可以比没有外婆照顾的妇女生育得更快,生下更多的孩子。更多的孩子意味着更健康,短的出生间隔迟早会在人群中占主导地位。 外婆假说可能非常有吸引力,并且具有科学可信度。尽管如此,在这一假设得到广泛接受之前,仍有一些问题需要回答。首先,在我们的进化史中,是否有足够多的女性拥有较长的生殖后寿命来增加适应?回顾第二章,历史上大部分时间的平均寿命都在35-40岁左右,超过60岁的人口不到2%。在没有外婆的家庭中出生的孩子将远远超过有外婆的家庭中的孩子。因此,选择压力——即改变个体遗传组成的事件(在本例中,是一位外婆提供护理)——对于一个能延长生殖后寿命或延长寿命的基因来说,可能不足以影响基因选择。 第二,年长的男性和女性兄弟姐妹是否有可能帮助照顾受抚养的儿童?例如,如果主要由年长的兄弟姐妹提供护理,那么长寿基因的选择压力不仅会很弱,而且根本不存在。考虑到史前人口中老年人的稀缺性,抚养子女的责任似乎更可能落在年长的兄弟姐妹而不是外婆身上。至少,照顾受抚养孩子的人会有相当大的差异。 最后,家庭结构和责任是什么?绝大多数早期社会都是父权制的。是男性而不是女性为孩子提供了最重要的照顾——收集食物。外婆可能为受抚养的孩子提供了情感、护理和耐心照顾,这是一项重要的工作,但与食物的生物必要性相比,这是微不足道的。如果没有男性为家庭提供食物,就不会有外婆作为假设的依据。 毫无疑问,在整个历史中,外婆在照顾外孙外孙女方面发挥了重要作用。甚至有可能外婆在进化史上对养育子女做出了更大的贡献,让她们的女儿生更多的孩子。更多的儿童是否能提高物种适应性,并使得生殖后寿命较长的适应性特征,还有待观察。支持外婆假说的数据既不广泛也不令人信服,几乎完全依赖相关分析,而非因果分析。然而,我们不应该马上否定任何至少存在一些支持性研究的假设。有了支持性的数学模型和高度受控的实验室研究,我们可能会发现外婆在家庭中的角色比做饭更重要。 |
随着时间的推移,魏斯曼意识到,他最初认为群体选择是其衰老理论的基础,这与达尔文关于自然选择作用于物种内个体间变异的观点相冲突。魏斯曼要么拒绝达尔文的自然选择理论,要么修改自己关于衰老进化基础的观点。他选择了后者。魏斯曼提出他的新理论是基于这样一种思想,即一旦某一特征对个体无用,自然选择就不再起到移除或维持该特征的作用。回想一下,魏斯曼认为生殖后阶段对生物体没有价值。由于大多数衰老的生理问题都发生在繁殖之后,衰老的特征既不会增加也不会降低适应性,因此衰老和/或衰老与自然选择的力量无关。魏斯曼将这些特征称为非适应性特征。衰老的中立性使魏斯曼保留了他的观点,即多细胞有机体的生殖后阶段是无用的,同时又保持了他严格的达尔文观点。
从达尔文关于变异、适应性的观点来看,中性或非适应性特征的出现往往会引起混淆。毫无疑问,这种困惑的产生是因为人们把自然选择作为进化的唯一力量。但就连达尔文也认识到,如果没有自然选择的力量,性状可能会变得“固定”:
(对繁殖)既不有用也不有害的变异不会受到自然选择的影响,要么像我们在某些多态性物种中看到的那样,成为波动因素,要么最终变得固定。(达尔文,1859年)
因此,衰老可能不会影响生殖。魏斯曼关于非适应性衰老的思想为预测衰老的中性特征理论奠定了基础,他认为,非适应性衰老要么是为生殖优势而固定的基因的副产品,要么是老年群体中基因的随机表达。
种群生物学家开发了逻辑方程来计算种群增长
孟德尔的遗传原理直到20世纪初才正式引入进化论。因此,达尔文和魏斯曼都无法将孟德尔的发现纳入他们的理论。孟德尔为达尔文自然选择的基本原理——变异是如何在一个物种中产生的问题提供了答案。根据孟德尔的说法,发生变异是因为每个父母传递给后代的特定基因的等位基因的形式可能略有不同。然后问题变成了,“这些等位基因在人群中占主导地位的速度和机制是什么?”对这些问题的研究催生了一种新的进化分析形式,即群体遗传学,这门科学专注于群体中等位基因变异的起源。
衰老和长寿的进化理论依赖于群体遗传学的一些基本原理,以更好地解释提高繁殖能力的等位基因在物种基因组中出现的频率如何影响群体的寿命。确定一个等位基因在种群中出现的速度需要对生殖潜力和种群增长有一个基本的了解——一个物种在最佳条件下自我繁殖的相对能力。
单细胞物种(如酿酒酵母)、简单多细胞生物(如秀丽隐杆线虫)以及培养基中生长的多细胞物种的细胞都被用来理解衰老和长寿的基本机制。尽管酿酒酵母、秀丽隐杆线虫和培养基中生长的细胞通过不同的机制繁殖,但它们的生长模式相似。这些增长模式对人口的寿命有重大影响。
简单生物种群的增长率反映了出生率减去死亡率,被称为内在自然增长率r。然而,所有简单和复杂物种的繁殖和生长都受到环境因素的限制,如食物、空间和温度,这会显著影响种群的增长和繁殖潜力。这些约束和其他约束统称为种群承载力K(环境容纳量),或因环境因素对种群规模施加的约束。Verhulst-Pearl logistic方程(方程3.1)中使用了这些概念来描述任何种群的种群增长,尤其是受缺乏流动性限制和/或在高度受控条件下维持的种群。
以下是verhulst-Pearl逻辑方程:
dN/dt=rN(K-N)/K (3.1)
此处
N=种群规模
r=内在自然增长率
K=种群环境容纳量
图3.3说明了对简单生物体的限制如何影响生长和繁殖以及影响基因选择。由于“母细胞”数量较少,培养初期细胞群增长缓慢。随着细胞数量的增加,因为有更多的“母细胞”来产生后代,所以生长速度也随之增加。因为食物充足,空间充足,所以这一时期的种群增长几乎完全由自然增长的内在速度驱动。随着食物供应和空间开始减少,繁殖率和种群增长都会放缓。最后,维持种群规模所需的空间和食物耗尽。如果不提供更多的食物和空间,细胞群就会衰老和死亡。当我们在第4章讨论复制衰老时,种群增长的逻辑方程变得很重要。
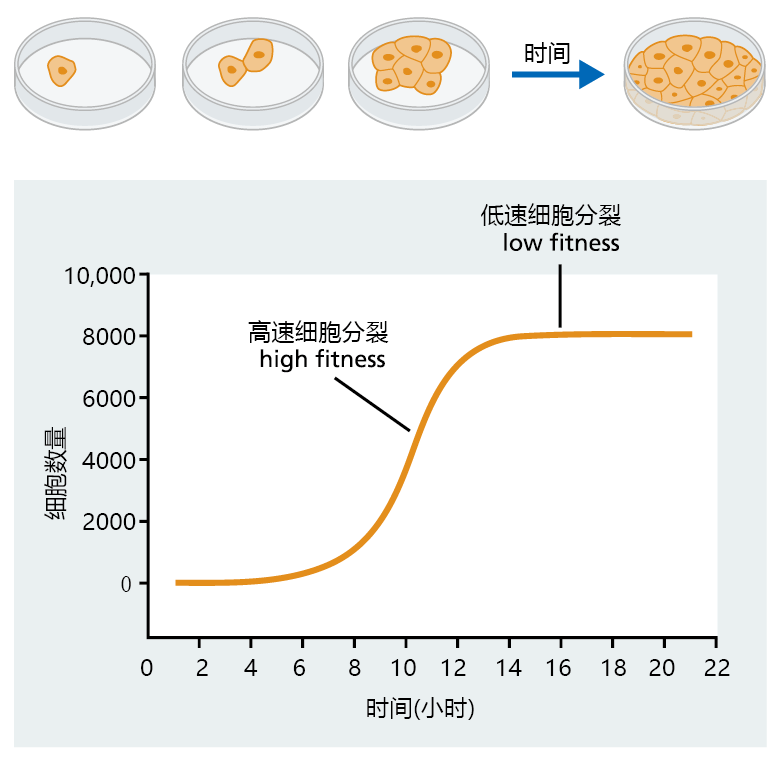
图3.3 Verhulst-Pearl 逻辑方程描述的假设细胞群的种群增长。在这个细胞群体中,对群体增长的限制由空间(容器的大小)和食物来表示。请注意,高适应期发生在培养物生命周期的早期,此时细胞分裂速度最大。当种群增长放缓并趋于平稳时,低适应期会在生命周期的后期发生。
这个简单的例子展示了群体遗传学的一个基本原理,它对老龄化和长寿的进化具有重要意义:高繁殖率发生的年龄具有最高的达尔文适应度,这仅仅是因为繁殖的个体数量。随着繁殖速度的减慢,适应性下降。因此,传递对生存和繁殖重要特征的等位基因比传递长寿的等位基因更容易被选择。这一重要概念有助于群体遗传学家形成关于选择生存的基因如何决定寿命的理论。
种群年龄结构描述复杂的真核生物中的达尔文适应度
更高级的真核生物不受环境条件的限制,其程度与我们前面的例子中描述的相同。复杂真核生物的运动性使动物能够寻找食物和水,在炎热的日子里找到荫凉处,在寒冷的日子里移动到隔离的地方。因此,对于复杂的真核生物来说,迁移意味着环境约束或K因子是可变的。在具有可变K因子的复杂真核生物中,确定等位基因固定的速率需要一组比用于简单生物体的逻辑方程更复杂的方程。这些方程描述了种群在生命周期内任何给定时间对后代的生殖贡献,或物种适应度,并构成了一种称为年龄结构分析的方法。
年龄结构分析可用于确定种群中的特定年龄组是否有助于自然选择对该种群的作用。它还可以用来回答这个问题:“一个人在什么年龄最有可能传递有利于物种生存的特征?”知道什么时候最有可能选择特定的等位基因,可以提供衰老、衰老和/或长寿是否是通过自然选择产生的信息。
在第二章中,我们介绍了使用生命表中的变量计算生存率(lx)时年龄结构分析的概念。作为适应的一个组成部分,生存预测在特定年龄组中有多少个体可以繁殖。因为在生殖开始后,每个连续年龄组的存活率都会下降,所以健康似乎也会下降。然而,存活率只是潜在健康状况的一个标志。为了计算人口的实际适应度,我们需要包括一个繁殖率的度量。这个指标被称为繁殖力,mx。
对于年龄结构分析,将种群分为年龄组。年龄分组过程没有固定的标准,但主要反映种群的生殖特征。周期性繁殖种群,例如每个季节只繁殖一次的动物(大多数鸟类和爬行动物),通常按繁殖季节的数量分组。繁殖持续发生的种群(许多哺乳动物)通常按合适的时间间隔分组,以天、月或年为单位。
繁殖率描述了繁殖种群的特定年龄适应性
周期性繁殖种群的年龄结构分析因繁殖季节中每隔一定的时间间隔产生后代而简化。代数方程可以用来计算总种群和繁殖季节的特定繁殖率。这些方程的数值结果称为净繁殖率(方程3.2)。种群的净繁殖率(R0)是所有繁殖季节的特定存活率(Lx)乘以特定繁殖的繁殖力(Mx)的积。
净繁殖率公式如下
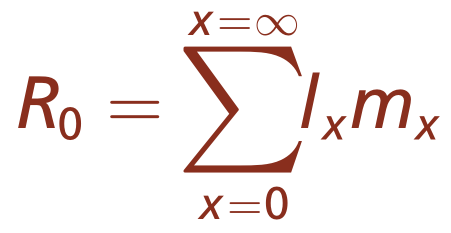
此处
R0=新生儿在其生命周期内可能产生的后代数量
lx=生存率,根据生命表计算
mx=繁殖力
净繁殖率提供了一个衡量繁殖能力的指标,因此也就是适应度。它最常用于周期性繁殖种群的分析。当净繁殖率应用于有繁殖季节的动物时,它被称为繁殖季节——特定繁殖率,表明哪些群体对生长和健康影响最大。例如,考虑图3.4中所示的假定的季节性繁殖种群。在每一个连续的繁殖季节,捕食和环境的恶化都会导致存活率下降。然而,种群中个体的性发育速度不同,导致在最初的几个繁殖季节繁殖力增加。在种群生命史的早期,繁殖力的增加将超过存活率的下降。在最初几个繁殖季节,存活率和繁殖力之间的反比关系导致繁殖季节的特定繁殖率增加。

图3.4 在一个假定的,均衡(R0 = 1)的季节繁殖种群使用净繁殖率进行的年龄结构分析。阴影部分代表种群规模。繁殖季节3为最高的繁殖季节(此时特定繁殖率为0.5),反映出繁殖力的增加。最年轻群体(繁殖季节1)和最长寿群体(繁殖季节6)动物的繁殖率最低(特定季节繁殖率为0)。也就是说,年轻个体的适应度最高,而年老的适应度最低。
随着存活率、繁殖力和繁殖率在每个连续繁殖季节下降,种群最终只剩下一小群不繁殖的长寿个体。再一次,自然选择倾向于提高生殖年龄存活率的等位基因,而不是增加物种寿命。
描述持续繁殖的物种种群的方程需要积分来确定增长率。统计学家阿尔弗雷德·洛特卡(Alfred Lotka)在18世纪瑞士数学家莱昂哈德·欧拉(Leonhard Euler)的工作基础上提出了描述持续繁殖种群中种群增长的方程式。因此,方程式中有两个名称(方程式3.3)。请注意,方程式3.3只是方程式3.2的积分,因此提供了关于生殖潜力和适应性的相同基本信息。
种群增长的Euler-Lotka方程的一般形式如下:
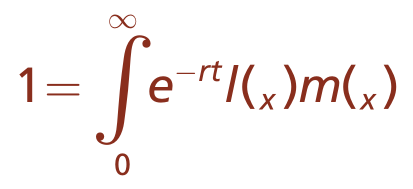
此处
e=数学常数
r=时间t时的种群内在增长率
l(x)=x时刻的存活率
m(x)=x时刻的繁殖力
Fisher描述了种群中生殖潜力和达尔文适应度之间的关系
Euler-Lotka方程为R.A.Fisher开发的数学模型奠定了基础,该模型可应用于衰老和寿命的进化理论。Fisher(1890–1962)第一个提出,在方程式3.3中求解r,即自然增长的内在速率,可以测量种群中个体的适合度。由于费舍尔的主要兴趣是预测性统计分析,他更关心当前的种群增长如何预测未来的个体繁殖力和适应性。为此费舍尔得出了一个衡量标准,即生殖值vx,他认为该值预测了个体未来对总种群繁殖贡献(方程式3.4)。
繁殖值,
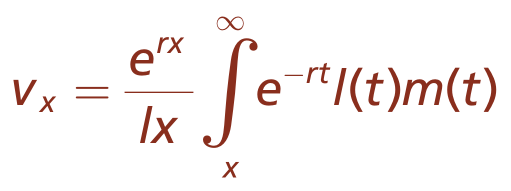
此处
vx=某个个体在x时刻的繁殖值
e=数学常数
r=种群内在增长率
lx=生存率
l(t)=在t时种群的剩余总生存率
m(t)=t时种群的剩余繁殖力
对于Fisher来说,vx回答了这个问题:“在持续繁殖的种群中,个体在什么年龄具有最大的适应性?”如图3.5所示,通过估计特定年龄组未来的生殖贡献,生殖值提供了潜在适合度的衡量标准。
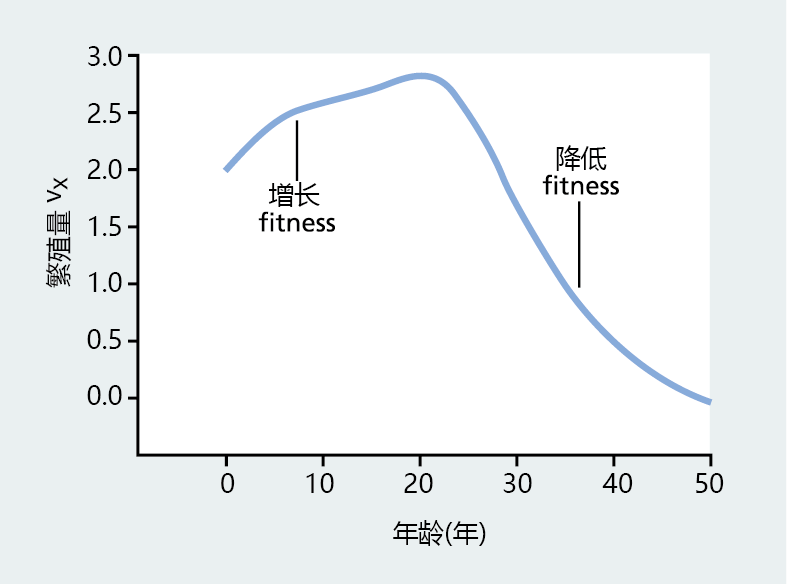
图3.5 1911年澳大利亚联邦女性生殖值(vx)。该图是通过在方程式3.4中插入记录的出生率和死亡率创建的。正如所料,生殖值在年轻时最高,随着人口老龄化而降低,接近生殖寿命的终点。请注意,vx的增加表示适应度的增加,vx的减少对应于适应度的降低。(改编自Fisher RA. 1930年,《自然选择的遗传学理论》,牛津:Clarendon出版社)
尽管Fisher帮助建立了长寿和衰老的进化模型,但他对生殖值和衰老之间关系的唯一暗示来自他的观察,即vx在死亡率开始增加的同时下降(图3.6)。他第一次提出,长寿可能是为生存到生殖年龄而固定的等位基因的副产品。Fisher的年龄结构分析在长寿进化中的更正式应用,必须等到Peter Medawar爵士的假说和W·D·Hamilton的数学,如下一节所述。
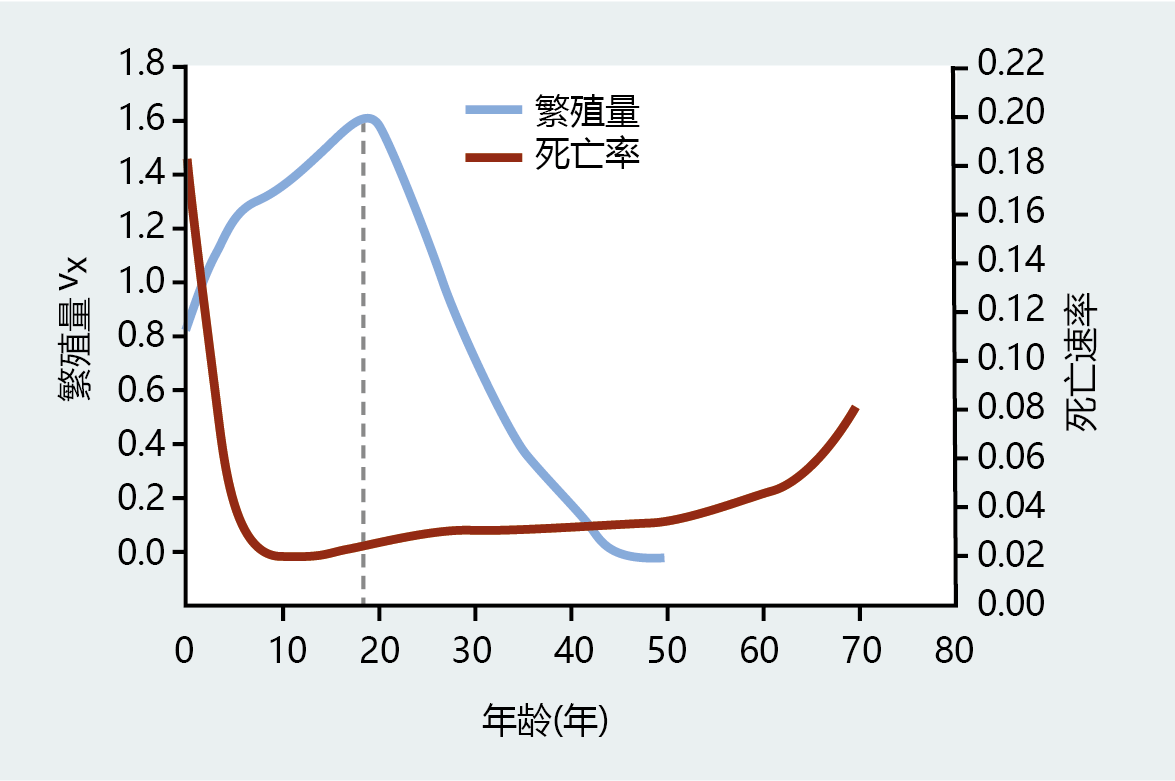
图3.6 1906年台湾女性的生殖值和死亡率。请注意,在虚线指示的点上,死亡率开始增加的时间与繁殖值vx开始减少的时间大致相同。(改编自Fisher RA. 1930年,《自然选择的遗传学理论》,牛津:Clarendon出版社)