
5.1 真核生物基因表达概述
从19世纪中叶到20世纪40年代中期,从达尔文开始,许多生物学家的共同研究工作使得基因被发现,基因是构建和维持生物的指令,位于真核生物细胞核内的染色体上。然而,对于这些基因是脱氧核糖核酸(DNA)的一部分还是蛋白质,仍存在一些争议。20世纪40年代末,随着DNA的结构越来越清晰,争议开始消散。罗莎琳德·富兰克林(Rosalind Franklin)和莫里斯·威尔金斯(Maurice Wilkins)进行的DNA X射线晶体学(一种显示分子中原子排列的方法)帮助詹姆斯·沃森(James Watson)和弗朗西斯·克里克(Francis Crick)更清楚地定义了DNA的结构,并表明DNA很容易重新排列,一定是基因的位置。1953年4月和5月,Watson和Crick发表了两篇论文,描述了DNA的结构以及四个核苷酸碱基腺嘌呤(A)与胸腺嘧啶(T)以及鸟嘌呤与胞嘧啶(C)的配对如何形成构建蛋白质的指令。
沃森和克里克的这些出版物描述了DNA的结构,彻底改变了生物学。在他们发现后的60年里,DNA中的信息如何转移到蛋白质中的氨基酸序列的基本机制已经被完全弄清楚了。在本章的第一节中,我们将探讨这个过程。
DNA转录产生互补RNA
基因表达开始于DNA在一个称为转录的过程中将其信息转移到核糖核酸(RNA)(图5.1)。从DNA模板合成RNA与您在第4章中了解的DNA复制过程有许多相似之处。双链、双螺旋DNA必须解旋以暴露碱基对。然后,一条(并且只有一条)DNA链充当RNA分子的模板,就像DNA复制一样,核苷酸一个接一个地添加。也就是说,RNA与DNA是互补的。然而,DNA复制和RNA转录之间存在一些显著差异。DNA中的腺嘌呤与碱基尿嘧啶(U)配对,而不是胸腺嘧啶,形成RNA中的碱基序列(图5.2)。也就是说,RNA中的四个碱基是腺嘌呤、尿嘧啶、胞嘧啶和鸟嘌呤。转录完成的RNA分子是一个单链分子。DNA复制和RNA转录之间的另一个重要区别是完成过程所需的时间和产生的分子数量。RNA分子的长度通常只有几千个核苷酸,相比之下,在中等大小的染色体中发现的2.5亿个碱基对的DNA。
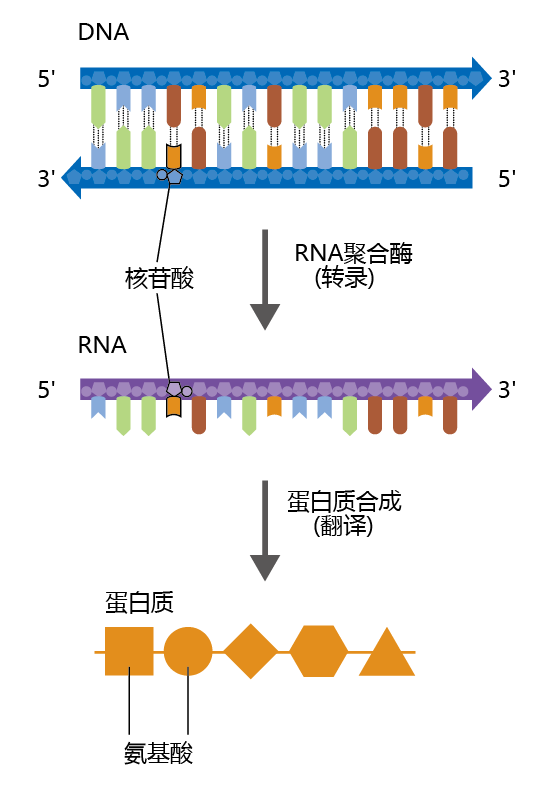
图5.1 转录和翻译。遗传信息从DNA转移到蛋白质。
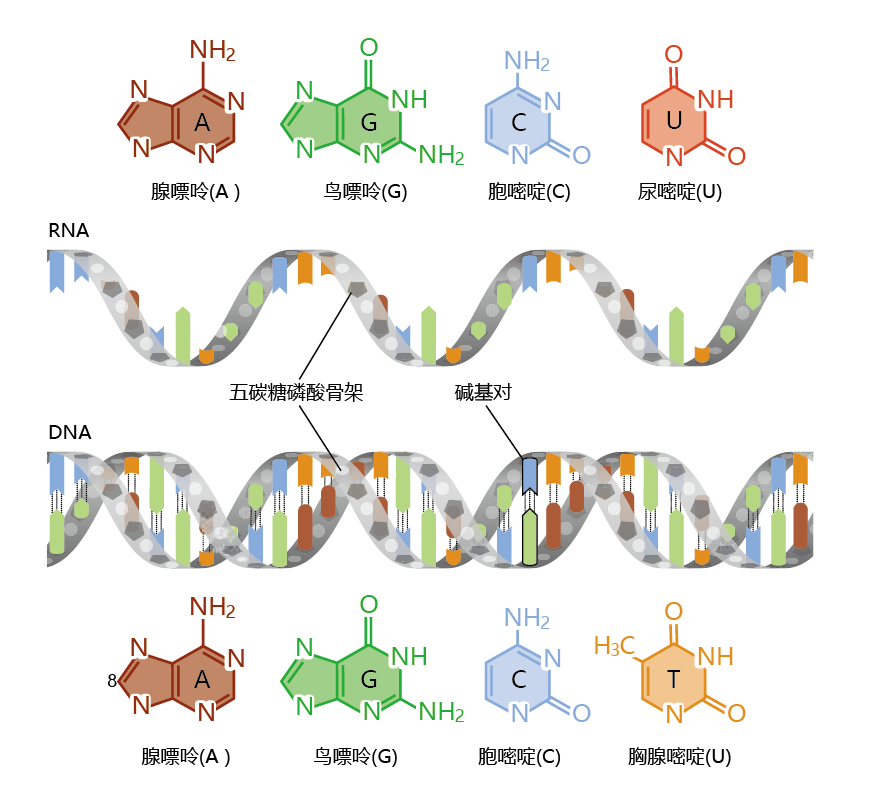
图5.2 RNA和DNA。RNA(顶部)和DNA(底部)由长链核苷酸组成,每个核苷酸由一个含氮碱基、一个糖和一个磷酸基团组成。DNA中的四个碱基是腺嘌呤、鸟嘌呤,胞嘧啶和胸腺嘧啶。RNA含有尿嘧啶而没有胸腺嘧啶。
DNA复制时间以小时为单位进行讨论,RNA转录以分钟为单位进行。RNA聚合酶(一种进行转录的酶)的几个分子可以同时作用于同一基因。当一个RNA聚合酶完成转录时,另一个开始。这允许在相对较短的时间内完成几个RNA转录,从而允许蛋白质的快速合成。
RNA聚合酶在识别DNA上的启动子区域时开始工作,这是一个特定的核苷酸序列,表明RNA合成的起点(图5.3)。单个RNA聚合酶在合成RNA时执行多种功能。首先,它打开双链DNA以暴露碱基。然后,酶的活性位点催化一种反应,将与DNA模板中的核苷酸互补的核苷酸添加到RNA中,一次添加一个核苷酸,在5′→3′方向。最后,RNA聚合酶将DNA倒带(rewind)成双螺旋结构。RNA转录产物继续延伸,当RNA聚合酶遇到DNA上另一个特定的核苷酸序列(即终止子)时,转录停止。

图5.3 DNA转录。RNA聚合酶与DNA的启动子区域结合。在启动过程中,RNA聚合酶解开两条DNA链并启动RNA合成。在延伸过程中,聚合酶继续组装RNA分子,其核苷酸序列与DNA模板链的核苷酸序列互补。当RNA聚合酶到达DNA上的终止位点时,它会与DNA模板解除结合,并释放新生成的RNA转录产物。
RNA聚合酶通过其特定形状识别DNA的启动子区域(图5.4),该区域是通过与一种称为通用转录因子的特殊蛋白质的启动子结合而形成的。普通转录因子通过定位一个称为TATA盒的序列来找到基因的启动子区域,因为它的序列主要由碱基胸腺嘧啶(T)和腺嘌呤(A)组成。普通转录因子与启动子区TATA盒的结合使得DNA向外弯曲。RNA聚合酶利用这种弯曲作为开始转录过程的标志。一旦RNA聚合酶与DNA结合并开始转录,普通转录因子就会被释放,再次用于另一个基因转录。
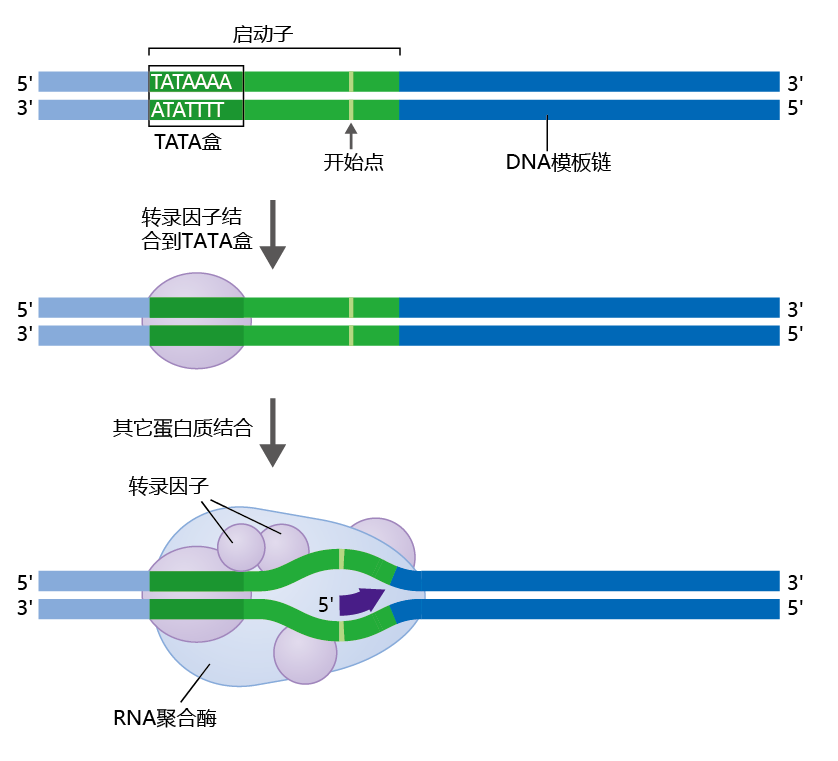
图5.4转录因子和转录起始复合物。转录因子蛋白与启动子区的TATA盒结合。转录因子介导RNA聚合酶的结合和转录的启动。
真核细胞在转录后修饰RNA
RNA转录产物包含基因的整个碱基序列,除了包含编码区的外显子之外,还包括被称为内含子的非编码区。由RNA转录产物剪接形成的另一种RNA,称为信使RNA(mRNA),是细胞用于合成蛋白质的RNA模板。mRNA只包含外显子,即基因的编码区。因此,在分子离开细胞核之前,必须从RNA转录产物中去除内含子。去除内含子的机制被称为RNA剪接(图5.5),这是由称为小核RNA(snRNA)的其他RNA分子进行的。snRNAs与其他蛋白质包装在一起,形成小核核糖核蛋白(snRNPs,发音为“snurups”)。snRNP与其他蛋白质结合形成剪接体,也就是进行RNA剪接的结构。剪接体切割内含子5′端的RNA转录产物(或前mRNA),如大多数内含子常见的几个短核苷酸序列所识别的。然后,snRNA沿着RNA向下滑动,直到它们在内含子的3′端找到另一组独特的核苷酸序列,然后在这里切割,将内含子从RNA中释放出来。然后将外显子的末端密封在一起。这个过程一直持续,直到所有内含子从RNA中去除,前mRNA变成mRNA。
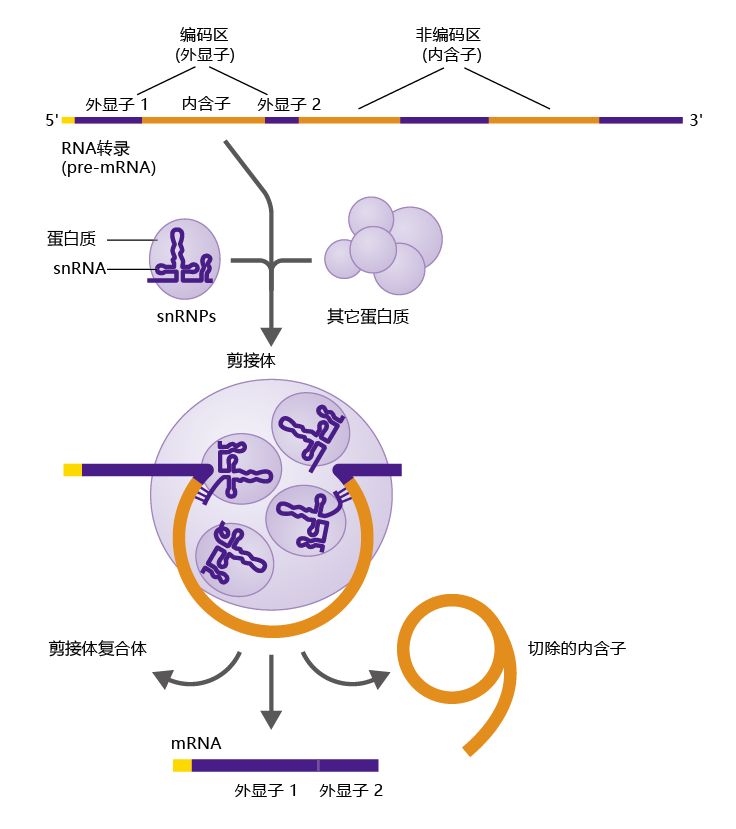
图5.5 RNA剪接。RNA转录产物与小核核糖核蛋白(snRNP)和其他蛋白质结合形成剪接体。剪接体切割内含子序列的两端,将其从转录产物中移除。外显子拼接在一起,形成信使RNA(mRNA)。
mRNA的合成发生在细胞核中,而翻译(由DNA编码的蛋白质的合成)是mRNA的主要目的,发生在细胞质中。因此,mRNA必须被运出细胞核。mRNA进入细胞质的运输对细胞来说是一个潜在的问题,因为剪接完成后,细胞核中存在许多RNA片段,如切除的内含子。换言之,细胞核如何只识别mRNA,然后只将这个分子从细胞核中运输出去?
核包膜包含称为核孔复合体的开口(图5.6)。这些孔允许具有特定结构的分子通过,这种结构是通过结合对mRNA具有特异性的蛋白质而产生的。例如,完整的和活性的mRNA包含独特的核苷酸区域,其特征是腺嘌呤碱基串,称为Poly-A尾。细胞核中普遍存在的聚A结合蛋白与这些区域结合,核孔复合体将这种复合体识别为mRNA。Poly-A结合蛋白只是细胞核中发现的几种仅与完整和活性mRNA结合的蛋白之一。一旦mRNA进入细胞质,结合蛋白就会被去除并降解,残留在细胞核中的RNA片段也会被降解。
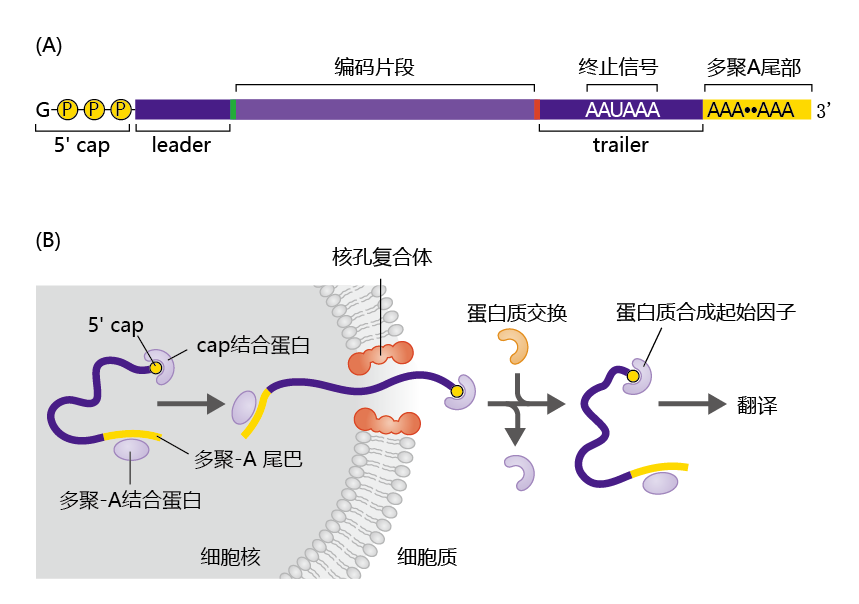
图5.6 转录后的RNA修饰。(A) 酶通过在5′端添加修饰的鸟苷帽和在3′端添加聚A尾来修饰mRNA分子的末端。(B) 两种蛋白质——一种连接到poly-A尾部的结合蛋白和一种结合到5′端的帽结合蛋白——与完整的mRNA分子形成复合物,使核孔复合物能够区分RNA片段和mRNA。一旦进入细胞质,帽结合蛋白被交换为蛋白质合成起始因子。
翻译是RNA指导的蛋白质合成
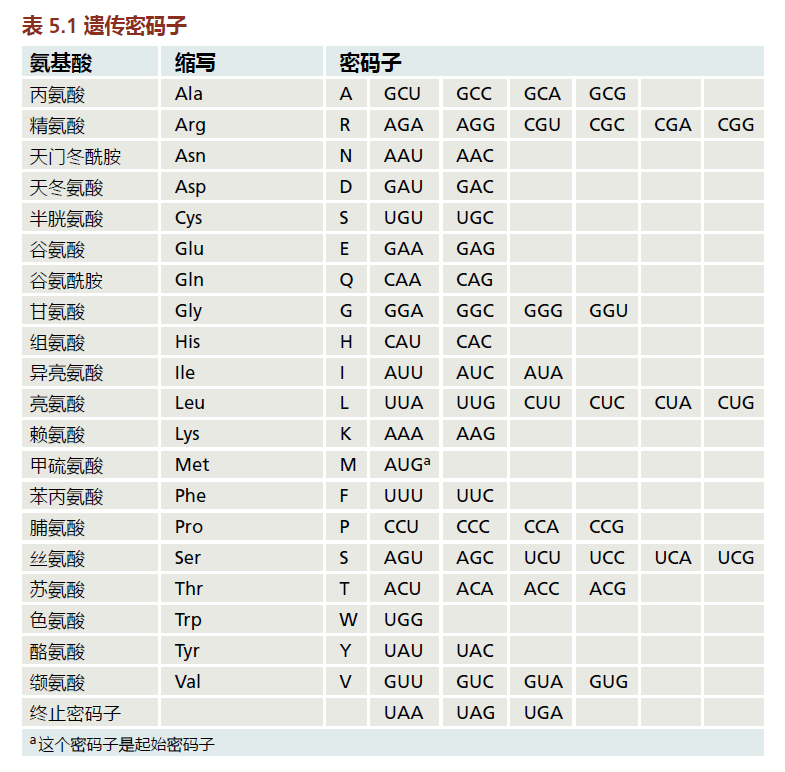
mRNA的核苷酸序列是用于蛋白质合成的编码。核苷酸以三个一组读取,称为密码子,每个密码子对应一个特定的氨基酸;密码子和氨基酸之间的这种对应关系称为遗传密码(表5.1)。一个氨基酸可以有一个以上的密码子;例如,密码子GCU、GCC、GCA和GCG都编码氨基酸丙氨酸(Ala)。密码子AUG编码的甲硫氨酸是起始密码子;三个密码子,UAA、UAG和UGA,被称为终止密码子,结束蛋白质编码信息。
mRNA不直接与氨基酸结合。相反,两种被称为衔接子转运RNA(tRNA)和氨酰基tRNA合成酶的特殊分子负责将mRNA中的核苷酸信息翻译成蛋白质的正确氨基酸序列的过程。tRNA是小分子,长约80个核苷酸。它们在一个位置上有一个与mRNA密码子连接的位点,在另一个位置有一个对应氨基酸特有的位点(图5.7)。氨酰基tRNA合成酶是一种将tRNA与其相应氨基酸共价偶联的酶。与氨基酸偶联的tRNA称为带电tRNA。
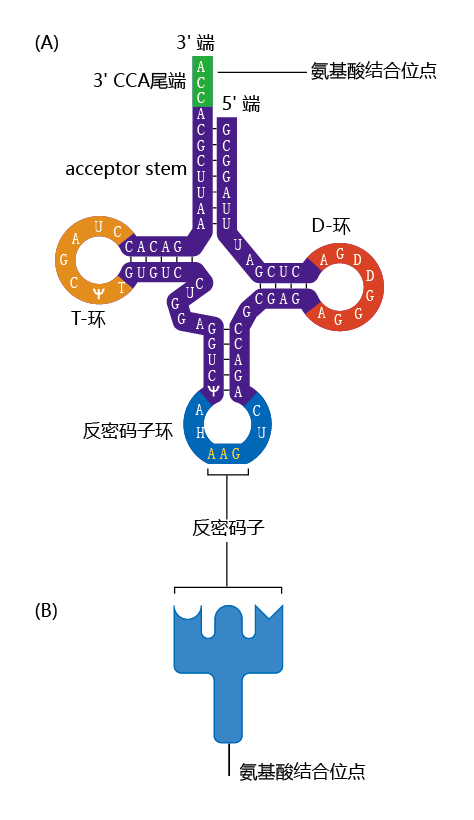
图5.7 tRNA的结构。(A) tRNA由形成多个双链区域的单链RNA组成,在这些双链区域RNA碱基片段相互配对。分子末端的单链环含有一个称为反密码子的三个碱基。另一端的短单链是氨基酸连接形成带电tRNA的位点。(B) tRNA分子。
到目前为止,我们已经发现mRNA含有蛋白质合成的遗传密码,tRNA和氨酰基tRNA合成酶一起将氨基酸带到mRNA中。将氨基酸结合在一起形成蛋白质的实际过程是一个复杂的多的过程,需要50-80种不同的蛋白质和将所有不同的分子(包括酶和其他蛋白质)保持在一起的物理位置。容纳所有这些蛋白质并为该过程提供空间的蛋白质制造复合物称为核糖体。每个细胞都含有数百万个核糖体,因此许多不同的蛋白质或同一蛋白质的多个拷贝可以同时快速合成(图5.8)。
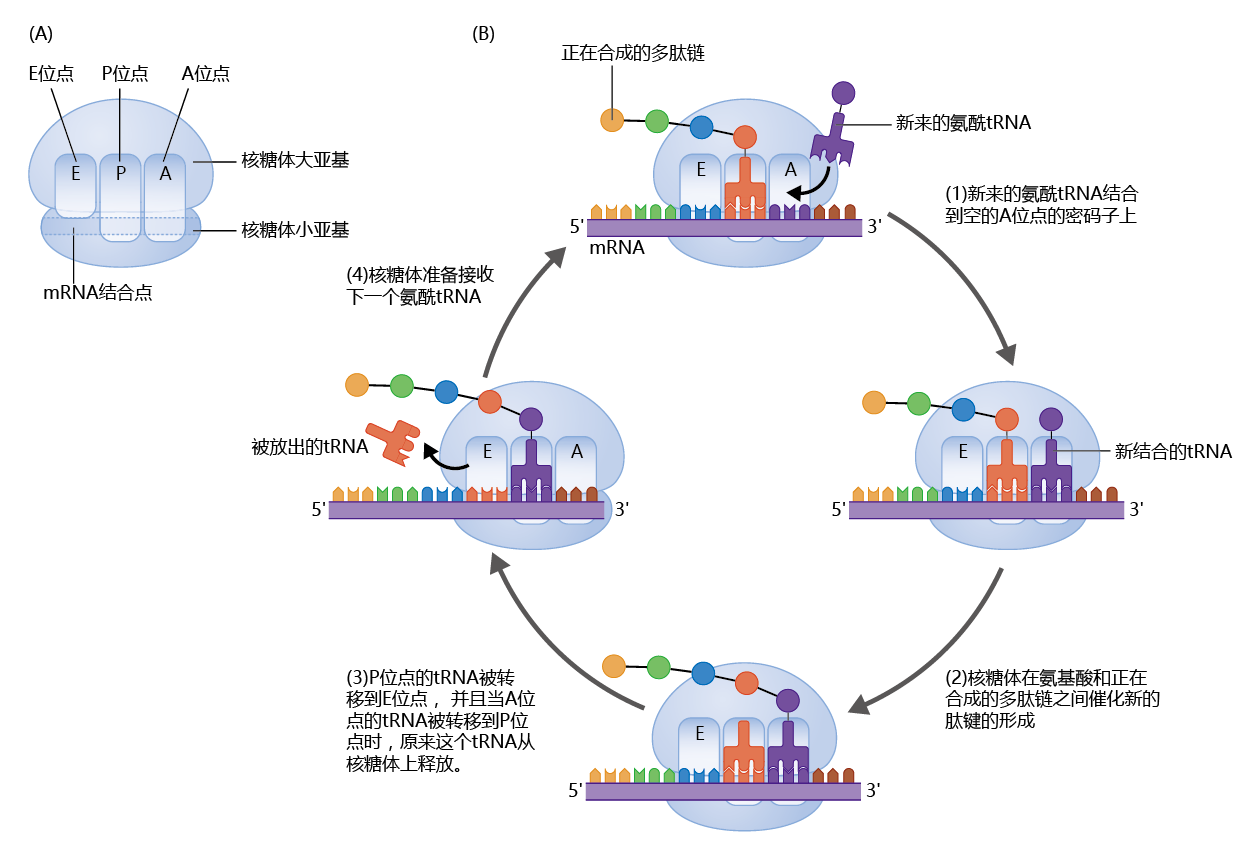
图5.8 核糖体上的蛋白质合成。(A)核糖体由两个亚基组成:大亚基和小亚基。核糖体有一个mRNA结合位点和三个tRNA结合位点,称为P、A和E位点。(B)翻译以四步循环进行。(1)进入的带电tRNA的反密码子与核糖体A位点的mRNA密码子配对。(2)带电的tRNA利用充电过程中产生的高能键的能量,将其氨基酸提供给生长中的蛋白质。(3)大亚基易位(如图所示向右),将tRNA分别从P和A位点移动到E和P位点,使A位点为下一个带电的tRNA打开。E位点的tRNA被酶从核糖体中去除。小亚基移动(向右)以匹配大亚基,拉动mRNA。(4)核糖体现在准备好在A位点接收下一个带电荷的tRNA,该过程再次从步骤1开始。这个循环继续进行,直到核糖体遇到mRNA中的终止密码子,此时完成的蛋白质从核糖体中释放出来。
翻译后蛋白质可以被修饰或降解
当核糖体释放完整的蛋白质时,蛋白质氨基酸之间的非共价相互作用导致蛋白质自身折叠,形成其三级结构。三级结构对蛋白质的活性状态至关重要。一些蛋白质需要在翻译过程后进行修饰才能完全激活。翻译后修饰有几种类型,但最常见的两种是磷酸化和糖基化,磷酸化是添加磷酸基团,糖基化是添加葡萄糖分子。通常,经过翻译后修饰的蛋白质被用作细胞内信使和信号。也就是说,它们通过一个非氨基酸分子来向其他蛋白质发出信号,这个非氨基酸分子可以与蛋白结合,也可以不结合。如本章稍后所述,DAF-16蛋白的翻译后修饰状态在秀丽隐杆线虫的寿命特征中起着重要作用。
一个功能正常、高效的细胞在很大程度上取决于其调节其组成蛋白的数量和结构的能力。真核细胞中已经进化出复杂的机制,以确保不正确折叠或受损的蛋白质被去除,并且单个蛋白质的浓度保持在最佳水平。这些机制涉及数千种参与数百种生化途径的蛋白质。在这里,我们简要讨论了大多数蛋白质降解途径的一些共性(图5.9)。
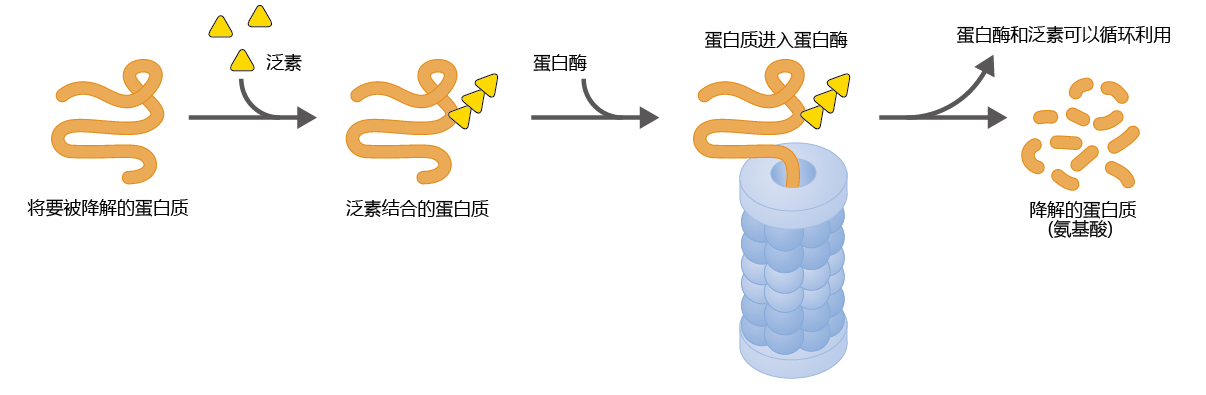
图5.9 蛋白酶体降解蛋白质。小的泛素蛋白附着在待降解的蛋白质上。蛋白酶体识别泛素标记(泛素化)蛋白,将其展开,并将其储存在它的中央腔中。蛋白酶体中的酶将蛋白质分解成小肽和氨基酸,然后释放回细胞质中。
一组被称为分子伴侣(chaperone)的调节性细胞质蛋白有助于维持其活性形式的蛋白质的正确三级结构。分子伴侣有两个主要功能:(1)它们帮助蛋白质正确折叠;(2)如果蛋白质没有正确折叠或受损,它们用另一种称为泛素的蛋白质标记其降解。一种被称为蛋白酶体的酶和其他蛋白质的大型复合物识别泛素标记的蛋白质。圆柱形的蛋白酶体含有被称为蛋白酶的酶,这些酶破坏蛋白质氨基酸之间的键(肽键)。这种分解释放的氨基酸返回细胞内氨基酸池,用于另一轮蛋白质合成。如第9章所述,涉及分子伴侣和泛素的蛋白质降解途径的功能障碍可能是阿尔茨海默病和帕金森病的潜在生化原因。