
10.3 MODULATING THE RATE OF AGING: PHYSICAL ACTIVITY
We are all familiar with the health benefits of regular physical activity. Adults of all ages who participate in regular aerobic activity have lower resting heart rates, lower blood pressure, lower levels of circulating triglycerides and LDL cholesterol (the “bad” type), and higher levels of HDL cholesterol (the “good” type) when compared with sedentary individuals of similar age (TABLE 10.1). The decrease in body fat that results from increased aerobic activity substantially lowers the risk of type 2 diabetes and certain types of cancer. Moreover, some recent research indicates that lifelong regular physical activity may be associated with a lower incidence of Alzheimer's and Parkinson's diseases.
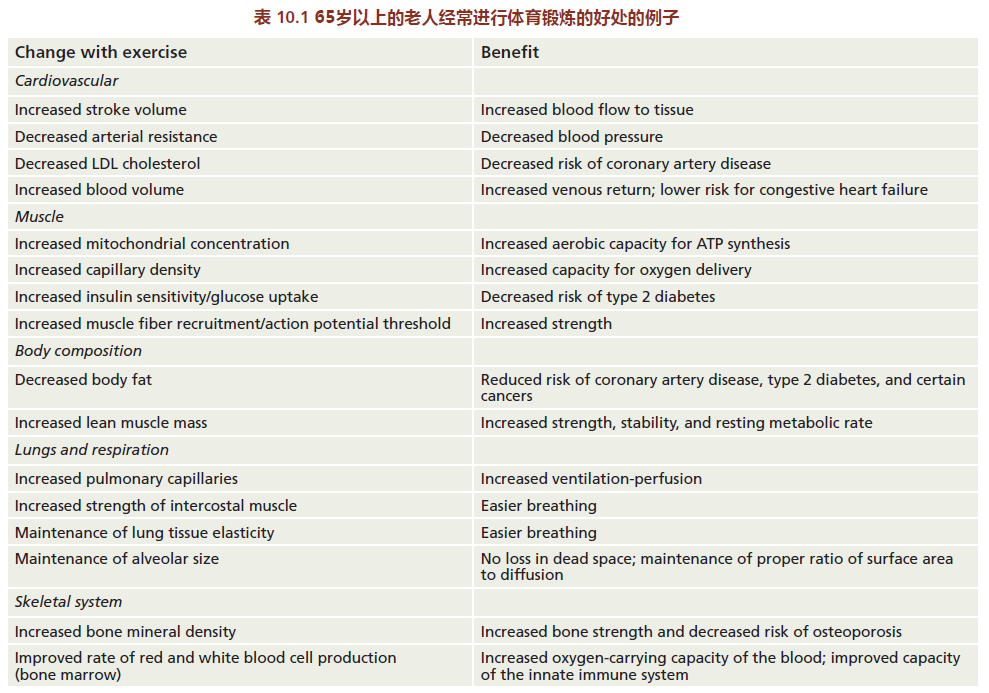
Regular physical activity lowers the risk of age-related disease, which, in turn, increases the mean life span. However, exercise does not increase maximum life span. This effect was shown in two classic studies published during the 1980s. In one study, the life spans of sedentary rats with different types of dietary restrictions were compared with the life spans of rats given access to running wheels. Rats with running wheels in their cages had mean (but not maximum) life spans slightly longer than those of animals without running wheels (Figure 10.7).
Interestingly, animals that were sedentary but calorie-restricted had the greatest mean and maximum life span. Thus, we can conclude that physical activity alters only the rate of aging, whereas calorie restriction alters both the rate of aging and longevity. Consistent with these findings are the human studies reporting that the length of the postreproductive life span directly correlates with the amount of regular physical activity. That is, the greater the amount of regular physical activity, the longer the post-reproductive life span and the longer the mean life span. In these studies, investigators showed that the increase in post-reproductive life span due to regular physical activity was most associated with a decrease in the risk for heart disease and cancer.
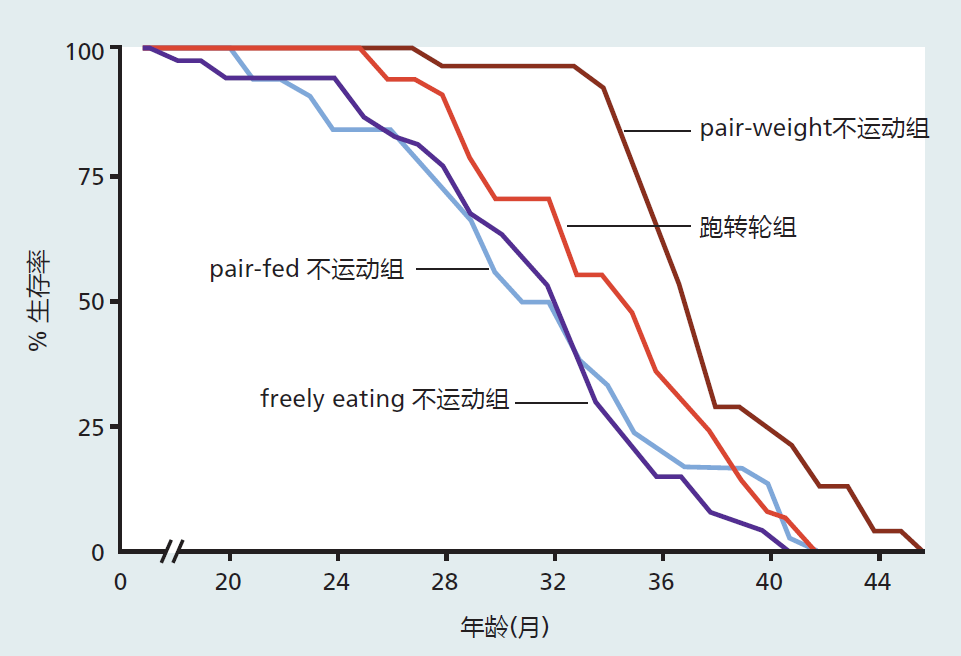
Figure 10.7 Survival curves for male rats with or without access to running wheels. “Pair-fed” sedentary rats received (daily) an amount of food equal to the amount eaten by the wheel runners. “Pair-weight” sedentary rats received an amount of food that resulted in a weight equal to that of the wheel runners; this amounted to ~30% of the CR diet. The wheel runners had a mean life span (34 months) greater than that of the pair-fed sedentary rats (31 months) and the freely eating sedentary rats (31 months) but less than that of the pair-weight sedentary rats (37 months). These data show that voluntary exercise (wheel running) significantly increases mean but not maximum life span when compared with sedentary animals. Calorie estriction increases both mean and maximum life span when compared with all other groups. In this section, we explore the biological basis for the increased function or maintenance of function resulting from regular physical activity. We begin with a brief introduction to the basic principles of exercise biology; adaptations that are thought to slow the rate of aging are discussed in terms of these basic principles. We end with some speculation on a general theory explaining why an intervention such as physical activity may slow the rate of aging.
10.3.1 Exercise increases the muscles' demand for oxygen
Most likely the first response you notice when beginning to exercise is the increase in your breathing rate, what pulmonary physiologists refer to as ventilation. The increase in ventilation occurs because chemoreceptors (neural receptors that bind specific chemicals) in the lungs detect increasing carbon dioxide levels in the blood returning from the extremities. Nerve impulses are then sent to the brain, which, in turn, sends signals back to the lungs, causing an increased rate of breathing. At the same time, the right ventricle of the heart senses a decrease in the amount of blood returning from the extremities, which is due to more capillaries opening up in the muscles. As a result, the heart beats faster to deliver more blood. All these physiological responses occur for one reason: the muscles need more oxygen to synthesize ATP so that they can carry out the increased work (rate of muscle contraction) of exercise.
Recall from Chapter 4 that the energy for contraction of muscle fibers comes from the breaking of one phosphate bond in ATP (ATP+H2O →ADP+ Pi+ energy+ heat). Because muscle contains only 5–10 seconds worth of stored energy, ATP must be synthesized continually during exercise. This process requires oxygen. In addition, the increased metabolic by-products of ATP synthesis, carbon dioxide and lactic acid, must be removed from the muscle cells to prevent acidosis, a decrease in pH that disrupts normal cellular chemical reactions.
ATP can be synthesized in muscle through two separate but connected pathways. The first, glycolysis, needs no oxygen (is anaerobic) and can use only glucose as the starting substrate for the synthesis of ATP. The anaerobic metabolism of glucose limits the amount of ATP that can be synthesized. For every molecule of glucose metabolized, only two molecules of ATP are synthesized. The other pathway of ATP synthesis, oxidative phosphorylation (discussed in Chapter 4), needs oxygen (is aerobic). Oxidative phosphorylation can utilize both fat and glucose to produce 16 times more ATP than glycolysis. The use of oxygen combined with the ability to utilize both fat and carbohydrates makes oxidative phosphorylation much more metabolically efficient than glycolysis. Humans use both systems simultaneously, at rest and during exercise, for ATP synthesis. At rest and during moderate exercise, the use of free fatty acids in oxidative phosphorylation predominates. When quick energy is needed, or during periods when ATP requirement outpaces a cell's ability to take up oxygen, glycolysis predominates.
Understanding how aerobic exercise stresses or overloads the oxidative phosphorylation system is the key to understanding how such exercise slows the rate of aging and leads to the benefits listed in Table 10.1. To this end, it is helpful to look at the physiological and cellular responses to a single bout of exercise. We'll use the example of a long-time sedentary individual who sets off to run a mile at increasing speed. In the first few moments, 5–10 seconds after she starts running, the cellular stores of ATP are used for muscle contraction(Figure 10.8). Because the intracellular stores of ATP are limited, the muscle cells need to supply more ATP to maintain the new level of muscle contraction. Even though it would be more efficient for the muscles to use oxidative phosphorylation for the synthesis of ATP, the body has not had enough time to recognize the increase in oxygen demand and to increase blood flow to the tissue. Thus, at this point, the needed ATP is produced by glycolysis, because this pathway does not need oxygen. However, the speed at which we can synthesize ATP in glycolysis comes at a cost. In addition to supplying only two ATP molecules for each molecule of glucose metabolized, glycolysis ends with the production of lactic acid. Humans are rather inefficient at neutralizing lactic acid, and if we continued to use only glycolysis for ATP synthesis, the muscle contraction would soon cease due to the drop in intracellular and blood pH.
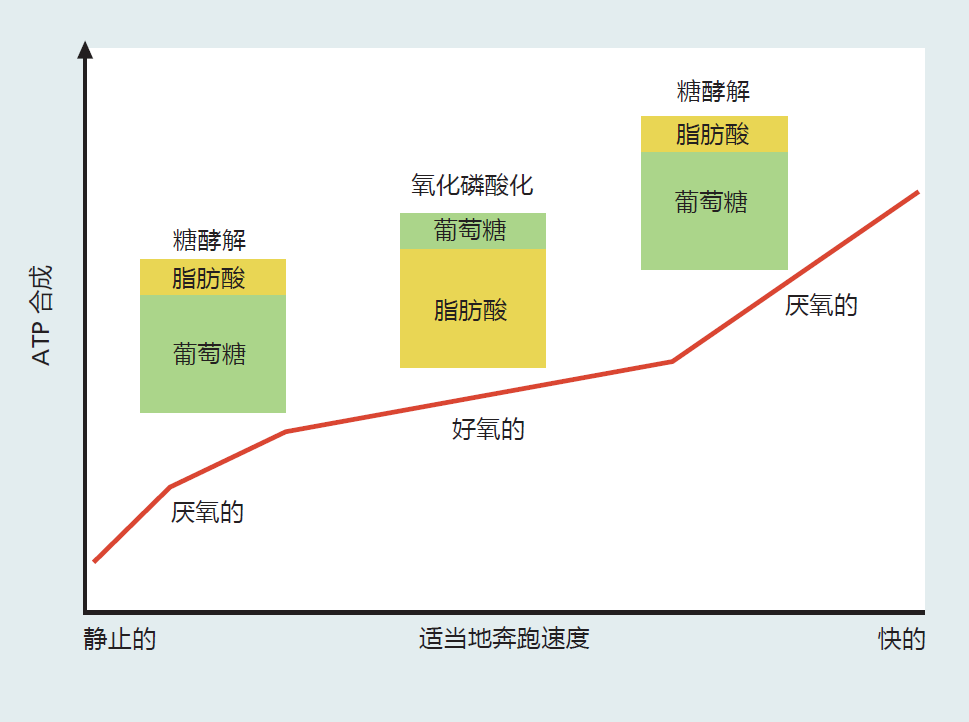
Figure 10.8 The metabolic pathways and energy substrates used for ATP synthesis during a single bout of exercise at different speeds.
After about 1–2 minutes of sustained moderate exercise (moderate meaning you can still talk while exercising), the body responds to the increase in muscle activity by increasing blood flow, which increases oxygen delivery. The increased supply of oxygen results in initiation of the oxidative phosphorylation pathway, a decrease in the reliance on glycolysis, and an increase in the use of fat as energy substrate. As long as this moderate level of exercise is maintained, ATP synthesis and exercise will continue without any negative metabolic consequences.
Now supposed this long-time sedentary individual decides to increase the speed at which she is exercising. The increase in speed increases her heart and breathing rate to meet the new demand for oxygen needed for ATP synthesis. She cannot increase her speed indefinitely, however, because the system has limits. Since she has been sedentary for a long time, her muscle cells have only enough metabolic capacity for a sedentary life. The heart and lungs are able to supply enough oxygen to the muscles, but the muscle cells do not have sufficient capacity to use the oxygen for ATP synthesis. This individual will eventually reach a point at which ATP synthesis through the oxidative pathway cannot keep up with the demand. At this point, the body starts to supply the majority of ATP through glycolysis.
10.3.2 Overloading cellular oxidative pathways increases the capacity for ATP synthesis
Our long-time sedentary individual has only enough aerobic metabolic capacity to maintain a muscle contraction rate consistent with a sedentary lifestyle. But, if she repeats the aerobic exercise daily over an extended period, she can increase her muscles' capacity for ATP synthesis. In every individual, aerobic exercise induces several physiological and cellular responses, such as increasing blood volume, red blood cell production, and expression of 蛋白质 and enzymes of the glycolytic and oxidative phosphorylation pathways (Figure 10.9). Repeating the aerobic exercise daily over a few weeks will make these responses permanent, even at rest. This metabolic adaptation explains the health benefits of exercise. As long as the ATP-synthesizing pathways are overloaded by aerobic exercise, the change will remain intact.
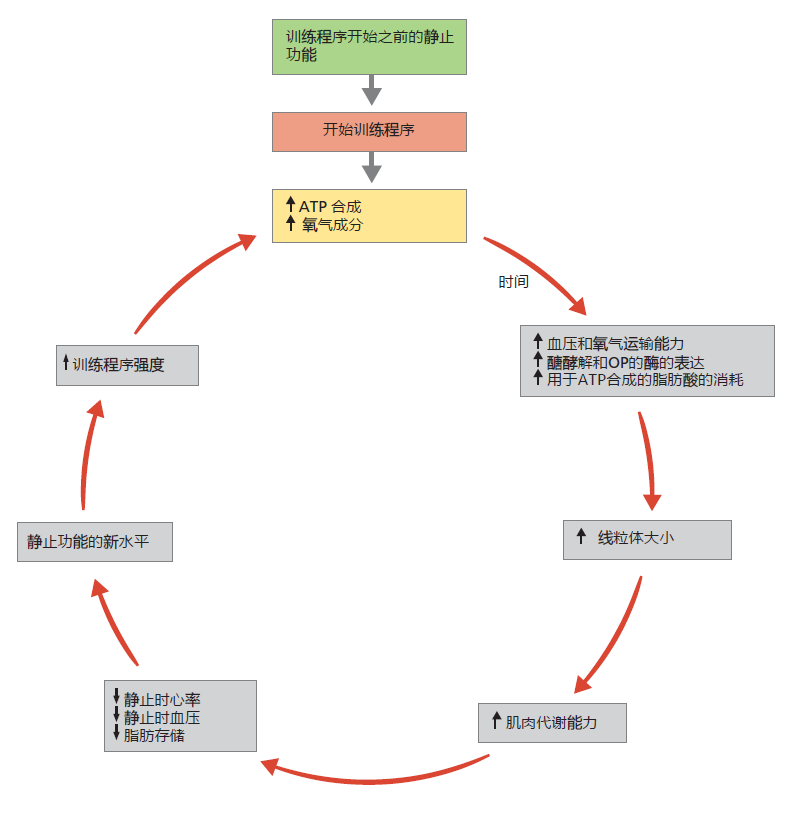
Figure 10.9 The training effect. The increased need for ATP synthesis during exercise is the driving force behind the benefits of regular physical activity. Increasing the capacity for ATP synthesis leads to improvements in cardiovascular function, with decreased resting heart rate and blood pressure, as well as a reduction of fat stores. Individuals of all ages can benefit from the training effect.
Increasing blood volume and red blood cell number through aerobic exercise increases the amount of oxygen per unit of blood delivered to the periphery. Moreover, because the aerobic exercise has increased the metabolic capacity of the muscles, oxygen extraction from the blood by the muscle cells has become more efficient. As a result, the heart does not have to pump as much blood to the periphery as it did before regular physical activity. The cardiovascular system adapts to this new level of muscle efficiency by reducing resting heart rate and blood pressure, thereby decreasing the work of the heart. In addition, a greater capacity for oxidative metabolism means a greater use of fat as energy substrate. Since fat is stored in adipose tissue, a regular physical exercise program will reduce weight, as long as there is no increase in dietary calories. Many studies have shown that reduction in weight lowers the risk of type 2 diabetes and coronary artery disease.
10.3.3 Regular physical activity prevents a decline in cellular reserve capacity
We presented the discussions on regular physical activity to illustrate that the biological process of aging can be slowed but not prevented. As you have seen, overloading a system, such as the capacity to synthesize ATP, results in adaptations that improve cardiovascular function, decrease body fat, and reduce the risk for certain diseases. In other words, a regular program of physical activity slows the rate at which systems fail due to increasing entropy—that is, it slows the rate of aging. These discussions have given you a brief introduction to how an intervention such as regular physical activity can slow the rate of aging. We now turn our attention to why: Why does physical activity slow the rate of age-related functional loss?
Regular physical activity slows the rate of aging because it helps to preserve reserve capacity in many physiological systems. Reserve, or redundant, capacity helps humans survive to reproductive age by providing a mechanism for defending against stress—such as disease, accident, or environmental insult—that exceeds the basal level of function. From the evolutionary viewpoint, the most successful hominids were those with the most reserve capacity that allowed them to reach reproductive age. But as we know, there would be no reproductive advantage to maintaining that reserve capacity after reaching reproductive age. At this age, reserve capacity would begin to decline.
The rate of decline in reserve capacity of a physiological system is determined, in large part, by the capacity that existed when the organism reached reproductive age. A higher reserve capacity results in a slower rate of aging. In Chapter 9, we discussed how, for women, reserve capacity of bone mineral content prior to menopause helps to prevent excess loss of bone mineral after menopause. However, many physiological systems retain the ability to increase reserve capacity throughout the life span. An increased reserve capacity will slow the rate of aging.
To illustrate how physical activity can slow the rate of aging, let's look at an example of reserve capacity in the human thermoregulatory system. The ability to maintain a constant internal temperature (37º C in humans) represents a survival advantage. Not having to rely on environmental conditions to raise or lower internal temperature means that humans can live virtually anywhere on the planet—a huge reproductive advantage in terms of obtaining sufficient food. Humans maintain a constant internal temperature through the heat produced in biochemical reactions. Basal levels of metabolism are sufficient to maintain the internal temperature when the environmental temperature, or ambient temperature, is between 20 º C and 25 º C. Ambient temperatures below or above this range require a physiological adjustment so that the internal temperature does not become too low or too high. For example, if the ambient temperature is 35 º C, we maintain our internal temperature by increasing blood flow to the skin, so that heat from the body is radiated to the atmosphere, and by sweating, transferring heat to the water in the blood, which is then released onto the skin for the cooling effect of evaporation. These mechanisms for heat dissipation rely on the reserve capacity of many physiological systems: heart rate increases to increase blood flow; stress hormones are secreted to signal the vascular system of the skin that internal temperature is rising; more skin capillaries open and larger vessels dilate. Maintaining internal temperature in a cold environment also requires responses from various physiological systems so that we conserve heat. Our ability to perform thermoregulation depends directly on the extent of our reserve capacity, and this reserve capacity is critical for survival.
Exercise increases our reserve capacity for defending our internal temperature against environmental challenges. Individuals who participate in regular physical activity maintain or increase the density of capillaries in muscle and skin, the rate of blood flow to the extremities, the stroke volume of the heart, and oxidative metabolic capacity. In other words, a physically fit individual should be able to respond faster and more efficiently to temperature challenges than a sedentary individual. Numerous studies have shown that aged people who participate in regular physical activity defend against a thermoregulatory challenge better than sedentary individuals.
Maintaining a regular program of physical activity has also been shown to increase or maintain the reserve capacity of the cardiovascular system and to slow the rate of aging in various physiological systems. These adaptations taking place during physical activity are being used to investigate the mechanisms underlying a human's ability to slow the rate of aging. Results from these investigations are still preliminary but are consistent with the hypothesis relating the maintenance of molecular fidelity to the rate of aging. For example, age-related atrophy of muscle tissue appears to be related to genes involved in the repair of damaged tissues. These genes are not expressed to the same degree in aged muscle as in young muscle. Aged muscle fibers seem to accumulate damaged 蛋白质, which, in turn, reduces molecular fidelity. Whether physical activity induces the expression of genes involved in the repair of damaged tissue remains to be seen. Nonetheless, physical activity may provide a suitable model to investigate the relationship between the maintenance of molecular fidelity and the rate of aging.