
3.3 TESTING 进化 MODELS OF 寿命
Thus far in our discussion, the possibility that longevity evolved has been supported by theoretical postulation and mathematically explicit computations. All theories, no matter how sound, must be put to the test in real life before complete acceptance is possible. To this end, several empirical investigations have been conducted to test the mathematical models of life-span 进化. In this section, we discuss results from a few investigations that link the 进化 of longevity to genes selected for reproduction.
3.3.1 Late-reproducing organisms have a lower rate of intrinsic mortality
Testing theories of 进化 can be challenging, given the length of time needed for natural selection to cause an adaptation. One laboratory method for modeling natural selection involves speeding up the process by collecting eggs (or offspring) from short-lived, rapidly reproducing species that have a particular trait. Eggs from animals without the trait of interest are discarded. The process continues for several generations until the trait has become dominant in the population. This testing process, called artificial selection, has been used extensively with Drosophila melanogaster to test the 进化 models of longevity.
Because artificial selection often does not involve manipulation of the 环境, this method is best suited to determining the intrinsic rate of mortality caused by genes currently present in the genome. In one such experiment, eggs were collected and separated into two groups: (1) eggs from flies that began reproduction early in their life span; and (2) eggs from flies that began reproduction late in their life span (Figure 3.11). After the two populations were established, the life span of each group was measured(Figure 3.12). The results were just as Fisher and Hamilton had predicted mathematically: flies that reproduced late in life lived significantly longer than flies in which fecundity was greatest soon after eclosion (emergence from the pupal casing). There did, indeed, appear to be a trade-off between fecundity and life span.
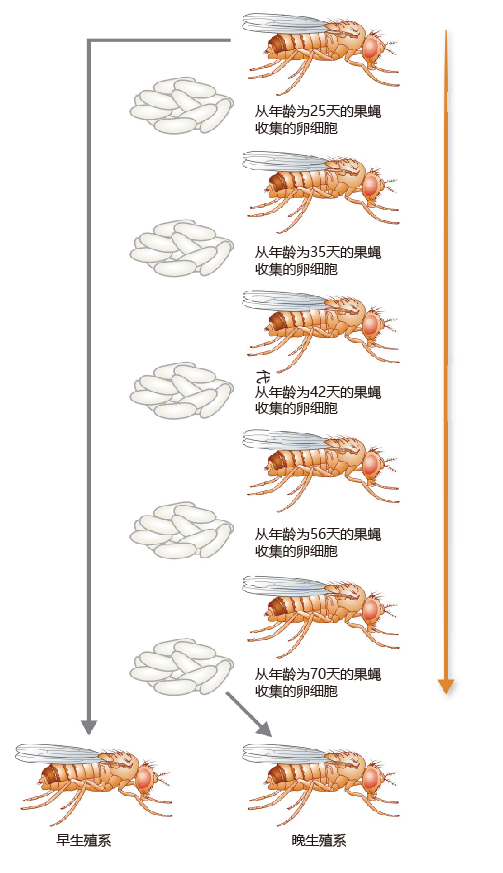
Figure 3.11 The use of artificial selection to create early- and late-reproduction lines in D. melanogaster. Eggs were collected and separated into two groups: eggs from flies that began reproduction early in their life span and eggs from flies that began reproduction late in their life span. Eggs from both groups were allowed to hatch, pass through their larval and pupal stages to produce adult flies, and begin reproduction. This process continued until two distinct populations of flies were formed, which took almost three years.
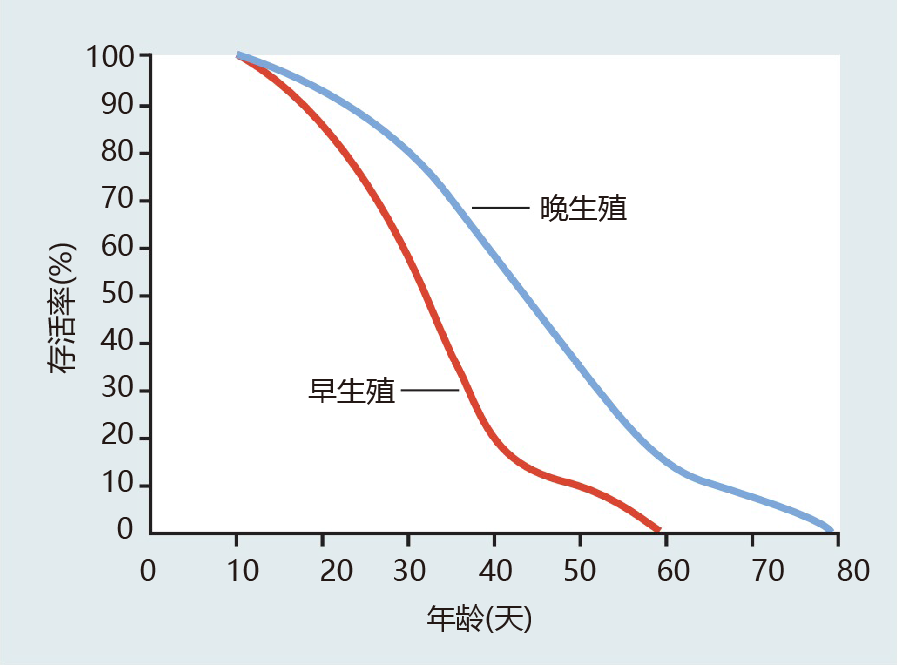
Figure 3.12 Survival curves for female D. melanogaster artificially selected for early and late reproduction. The early-reproduction line (red) has an average age at highest fecundity of 4–5 days, and the late-reproduction line (blue) has an average age at highest fecundity of 8–12 days. These results demonstrate that genes selected for their timing at the start of reproduction also affect the length of the life span. (Adapted from M.R. Rose, 进化 35:1004–1010, 1984. With permission from Wiley-Blackwell.)
3.3.2 Genetic drift links life span to reproduction
The artificial selection experiment described above eloquently demonstrates that longevity evolved and may be linked to genes selected for reproduction. However, artificial selection does not test whether or not 环境al conditions, the primary driving force of natural selection, can work to fix genes that extend the life span. To do this, we need to see whether there is a similar trade-off between reproduction and life span when extrinsic factors, such as differences in predation and starvation, are introduced. This can be tested under laboratory conditions. For example, Sterns and colleagues allowed eggs from two groups of flies to hatch and progress through the larval and pupal stages into adulthood. Prior to the start of reproduction, 90% of the adults in one group are killed, a method simulating high predation. In the other group, the density of flies in the housing chamber is reduced to a level at which all flies have sufficient food and no predation throughout reproduction, a method simulating 环境al conditions leading to a stable population. 进化ary theory predicts that flies subject to high predation would reproduce early and have more offspring than flies reared in a stable population without predation. And, indeed, this prediction was supported by experimental data from this type of experiment. In other words, the methods of this experiment simulated natural selection; flies exposed to high or low predation adapted to their 环境.
Of course, we are interested in whether or not genes selected for a particular reproductive scheme can determine the length of life. 进化ary models of longevity predict that flies with early reproduction (the high-predation group) should have a faster rate of mortality than late-reproduction flies (the low-predation, stable group). Again, this prediction was supported by the data (Figure 3.13). Thus, both the intrinsic and extrinsic rates of mortality are linked to the timing of reproduction, which, in turn, determines the length of the life span. The mathematical and verbal predictions of Fisher, Medawar, and Hamilton are supported.
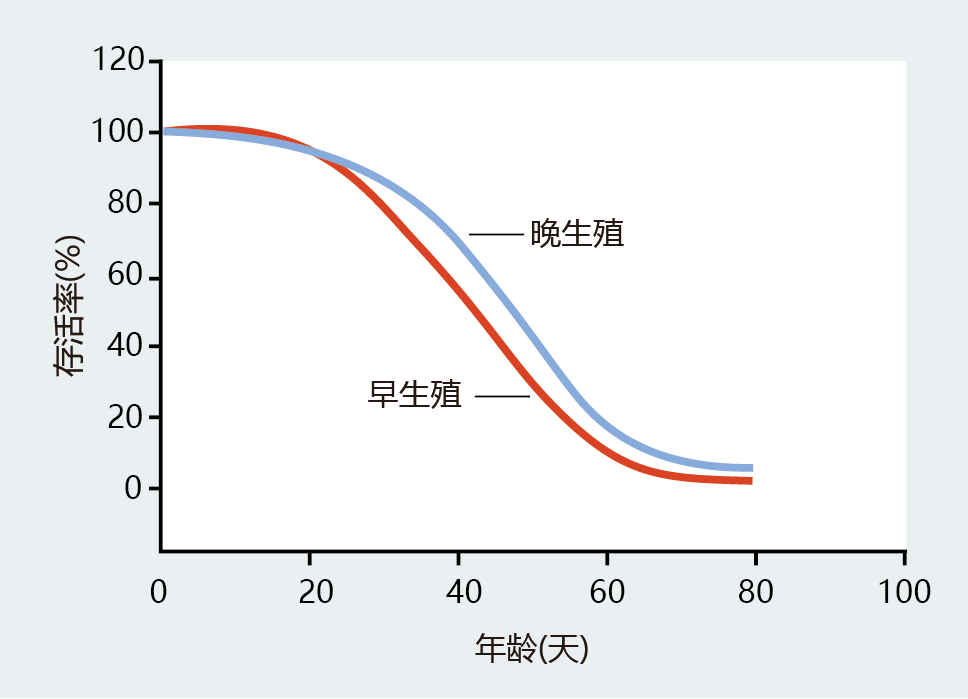
Figure 3.13 Relationship between extrinsic mortality rate and duration of life span. As described in the text, flies are subject to controlled 环境al conditions. Simulation of high predation leads to the development of early reproducing flies, while simulation of low to no predation results in a line of flies with late reproduction. Data collected from such experiments show that late-reproducing flies have slightly longer mean and maximum life spans.
It is important to demonstrate the trade-off between reproduction and life span in wild populations, not just in laboratory experiments. Investigations in wild populations compare the reproduction schedule and life span of a single species living in two different 环境s. For example, turtles living on an island with low predation can be compared with the same species living on the mainland with high predation. In most cases, populations with high predation (high extrinsic mortality) have significantly shorter life spans than populations of the same species living in areas with low predation (low extrinsic mortality). This finding has held for several different land-based phyla, including birds, mammals, and reptiles. However, similar studies in fish have not supported the trade-off hypothesis. The different result in fish than in land-based phyla demonstrates that theories, while supported by laboratory tests, may not always follow the prediction in the wild. Nonetheless, there is enough evidence from the study of wild populations to generally support the 进化 basis of longevity.
3.3.3 Results from testing the 进化ary theory of 寿命 have changed research in 衰老生物学
Before laboratory testing of the 进化ary theory of longevity, biogerontology was an observational science lacking clear and precise definitions of aging and longevity (see the discussion in Chapter 1). Without these critical definitions, identification of biological mechanisms underlying aging and longevity was difficult, due in part to researchers defining aging and/or longevity to fit their hypotheses, rather than the other way around. Moreover, biogerontology lacked consensus as to the direction of research. The results from laboratory experiments on the 进化 of longevity described here separated the biological processes of aging and longevity; more precise definitions were now possible. In addition, the understanding that longevity evolved through genes selected for reproductive success and that aging reflects unregulated, random, and stochastic mechanisms led to a much clearer research direction for biogerontology.
Separation of the 进化ary origins of aging and longevity revolutionized research in biogerontology. Biogerontologists were now able to formulate hypotheses based on an accurate description of how a species' longevity became part of its overall life history. To this end, geneticists looking for genes affecting longevity could narrow their search to genes that connect longevity to the reproduction schedule for the species. In Chapter 5, you‘ll learn that many genes have been identified that have dual effects—that is, they were selected for reproductive success but also affect longevity. And in Chapter 7, you'll learn that results from the laboratory testing of the 进化ary theory of longevity established a solid foundation for, and helped to give rise to, the study of gerontological biodemography, a science that integrates biological knowledge with demographic research on human longevity and survival.