
4.4 细胞衰老的原因:受损生物分子的积累
在第一章中,我们强调了衰老原因和衰老机制之间的区别,前者只能有一个,后者被认为是无限的。我们现在详细描述衰老的根本原因。不管细胞经历了什么样的衰老机制,人们几乎普遍认为细胞衰老是受损蛋白质在细胞内积累的结果。因此,在本节中,我们将探讨生物分子损伤积累的方式和原因,以及这种损伤可能导致衰老速率改变的原因。
生物分子服从热力学定律
与宇宙中的所有物质一样,生物分子也受制于热力学定律,即控制功、能和热之间关系的物理定律。热力学第一定律适用于能量从一种形式到另一种形式的转换。将能量从一种形式转换为另一种形式时,转换前后系统的总能量保持不变。也就是说,能量既没有被创造也没有被破坏。在能量从一种形式转换到另一种形式的过程中,我们遇到了热力学第二定律:能量从一种形式转换到另一种形式的效率不是100%,一些能量变得无法使用。除非系统接收到新的可用能量输入,以取代作为热量损失的不可用能量,否则系统将随机重新排列,也就是说,它将朝无序(熵)的方向移动。这种增加熵的运动会导致受损蛋白质的积累,从而导致衰老。
生物过程的总能量叫做焓(H),它等于可用能(自由能,G)加上不可用能(熵,S)。方程4.1描述了这种关系。
系统的总能量如下所示:
H=G+TS (4.1)
此处
H=焓,系统中的总能量
G=可用能量或自由能
S=不可用能量或熵
T=系统温度
因为能量存在于分子的键中,H、G和S不能在活体生物系统中直接测量。然而,只要我们知道反应发生的温度,我们就可以测量每个量的变化,用希腊字母δ(Δ)表示。我们首先确定一个反应是释放还是消耗可用能量。也就是说,ΔG=G产物−G反应物。如果反应增加了可用能,ΔG的符号为正;如果反应降低了可用能,ΔG的符号为负。因为第一定律告诉我们,一个反应的总能量是不变的,所以G的任何变化都必须伴随着H和S的相等和相反的变化,但H不能被测量。因此,我们可以测量ΔG和ΔS,并求解ΔH(方程式4.2)。
化学反应的可用能量变化如下:
∆G=∆H− T∆S(4.2)
此处
ΔG=反应过程中自由能的变化(G产物−G反应物)
ΔH=系统添加或释放的总能量
ΔS=熵的变化
T=系统温度
生命需要不断维持秩序和自由能
生物过程在反应中使用具有大量可用能量(高度有序)的分子,从而使得生物体生殖、生长和修复。在这些反应中,由于第二定律,不可用的能量被释放,这降低了进行维持生命的额外生化反应所需的可用能量。根据第一定律,随着可用能量的减少,系统的熵和无序度(或不可用能量)肯定会增加。为了恢复秩序,从而维持生命,生物体必须不断提供新的可用能量,以取代因熵而损失的能量。肌肉的收缩说明了生物过程中的热力学原理。
肌肉细胞(肌肉纤维)将ATP的化学能转化为称为收缩的机械能(图4.14)。ATP转化为二磷酸腺苷(ADP)+Pi(无机磷酸盐)释放可用能量,驱动收缩。由于热力学第二定律,以热的形式释放出不可用的能量。事实上,断开磷酸盐高能键产生的80%的能量是以热的形式释放的;随着肌肉纤维的每一次收缩,熵和无序度增加,可用能量减少。如果该系统提供了一种机制,可以替代因熵而损失的可用能量,那么肌肉可以放松,为下一次收缩做准备。也就是说,随着ADP转化为ATP(也就是说,更多的键,更多的可用能量,更多的阶数),失去了熵的阶数得以恢复,如图4.14所示。相反,如果失去的能量无法恢复,肌肉纤维将保持收缩状态,导致缺氧,细胞将开始死亡。如果没有额外的自由能,也就是说,如果我们死了,热力学定律就会完成,肌肉纤维就会达到能量平衡。
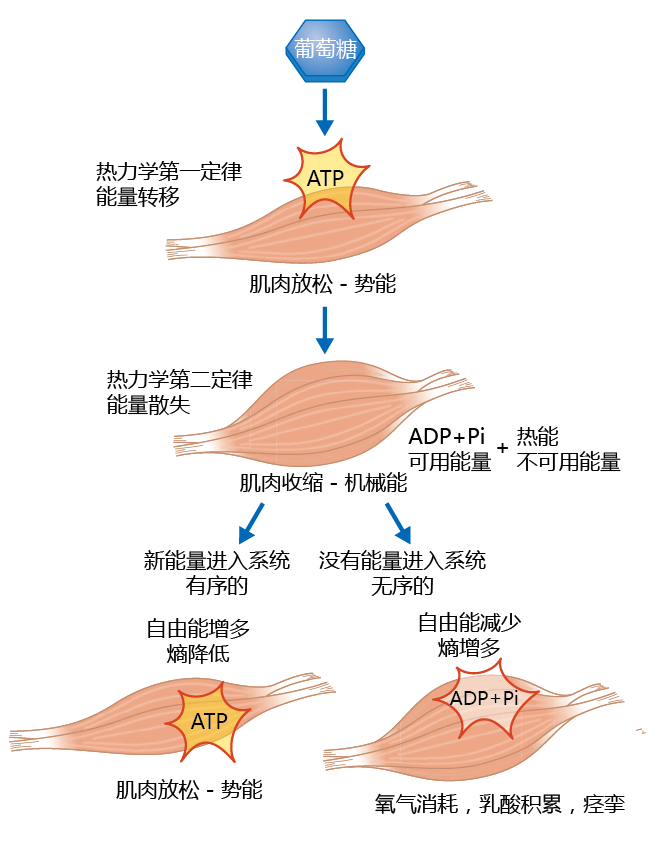
图4.14 应用于肌肉细胞的热力学第一定律和第二定律。根据热力学第一定律,肌肉细胞将葡萄糖能量转化为ATP能量,为收缩做准备。ATP转化为ADP+Pi释放出可用于促进收缩的能量。由于热力学第二定律,热量也会释放出来。如果更多的葡萄糖进入系统,取代因收缩而损失的能量,肌肉就会放松,为下一次收缩做准备。如果没有葡萄糖可以转化为ATP,肌肉将保持收缩状态。
衰老的机制是分子保真度的丧失
回想一下,分子的原子结构决定了它的功能。我们把这个原理称为结构-功能关系。如果一种蛋白质中哪怕有一个氨基酸的序列不正确,那么该蛋白质的生物活性就会降低。维持分子的适当结构,即分子保真度,对生物体的生存至关重要,需要持续输入可用能量。换句话说,生物体不断与熵的增加作斗争,而成功的生物体则是那些通过保持分子保真度而赢得战斗的生物体。
为了恢复因熵而损失的自由能并维持秩序,生物体需要投入大量资源才能存活到生殖年龄。在最基本的层面上,成功存活到生殖年龄仅仅反映了被选择来维持蛋白质分子保真度的基因。然而,正如你在第三章中所了解到的,在达到生育年龄后,维持这种高水平的资源投资将没有生育优势。宇宙的法则最终将成为规则,缓慢走向能量平衡将导致分子保真度的丧失、细胞的无序,以及由于受损蛋白质的积累而导致细胞功能的降低。细胞衰老反映了由宇宙基本定律——熵增无序——导致的受损蛋白质的积累。
衰老反映了受损生物分子在细胞内的积累
现在有压倒性的证据表明,时间的推移会导致分子结构的变化。虽然这些变化背后的机制尚不清楚,但我们可以自信地说,在性成熟后,受损生物分子的细胞积累“加速”。由于结构决定功能,较老的细胞在执行正常操作时效率较低。我们可以用维持分子保真度(有序)和熵(无序)之间的平衡来看待体细胞衰老(图4.15)。由于自然选择的力量,在发育过程中,高功能细胞至关重要的时期,基因被选择来维持分子保真度。根据一次性躯体理论(见第3章),随着时间的推移,外在的衰老速度会对生物分子造成损害。因为有助于维持分子保真度的蛋白质也受第二定律的约束,修复或替换受损生物分子的能力下降,导致受损生物分子在细胞内随时间累积。当熵开始超过维持分子保真度的速度时,细胞就不能再维持正常功能,细胞就会死亡。
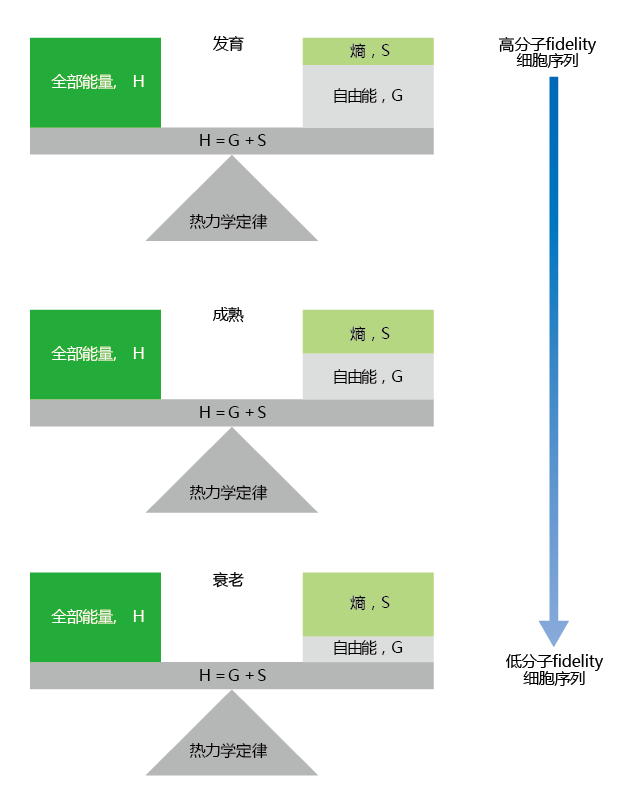
图4.15 分子保真度和能量平衡。分子保真度和细胞秩序在生殖年龄(发育)前的阶段得以维持。可用能量的替换超过了熵损失的能量。随着我们年龄的增长超过了生殖(成熟)的开始,熵的增加会导致分子保真度的损失和受损蛋白质的积累,从而导致细胞紊乱。衰老的特点是分子保真度低,细胞无序程度增加,因为熵的增加速度快于新的可用能量的引入。
我们故意模糊了可能导致细胞衰老的分子去保真度的确切损伤类型。其原因是,细胞损伤的确切类型对衰老过程基本不重要。损伤的类型和数量在不同的细胞类型之间有很大的差异,没有一种损伤过程可以被认为是细胞衰老的主要原因。例如,在本章中,我们使用以氧为中心的自由基造成的损伤来描述正常新陈代谢中的随机事件如何导致损伤的累积和衰老的出现。其他章节描述了向蛋白质中添加葡萄糖(称为糖基化)如何改变蛋白质的结构并导致损伤的累积。在第9章中,你将了解到蛋白质的错误折叠可能会导致不溶性聚集体,这可能是神经疾病的前兆。与糖基化蛋白质或不溶性蛋白质聚集体相比,很难证明以氧为中心自由基引起的衰老相关损伤对衰老过程更重要。
关于损伤累积原因的讨论中要传达的重要信息不是损伤累积的类型,而是细胞损伤是根据宇宙的基本规律发生的。因此,与我们对物质世界中任何其他物体的控制相比,我们无法控制人体内衰老的恶化影响。我们可能能够阻止一种类型的损害,但却发现另一种类型的损害已经开始。这将持续到细胞屈服于第二定律。当一个或多个特定器官的足够多的细胞由于分子去保真度而停止运转时,生物就会死亡。你在第10章中了解到,我们可以通过各种方式减缓衰老的速度,活得更长、更健康,但衰老最终会占上风,宇宙会夺走我们的生命。