
5.5 秀丽隐杆线虫寿命的遗传调控
秀丽隐杆线虫只有1毫米长,生活在几个气候区的土壤中;30多年来,它一直被用作长寿遗传学的模式生物。这种蛔虫主要以细菌为食,但在实验室中,它可以在几种不同的生长介质上生长。
秀丽隐杆线虫有两种性别,雌雄同体和雄性。雌雄同体同时产生精子和卵子,并通过自我受精繁殖(图5.35),而雄性只产生精子。雄性以极低的频率(1/500)自发出现,并能使雌雄同体受精;雌雄同体不能使其他雌雄同体受精。
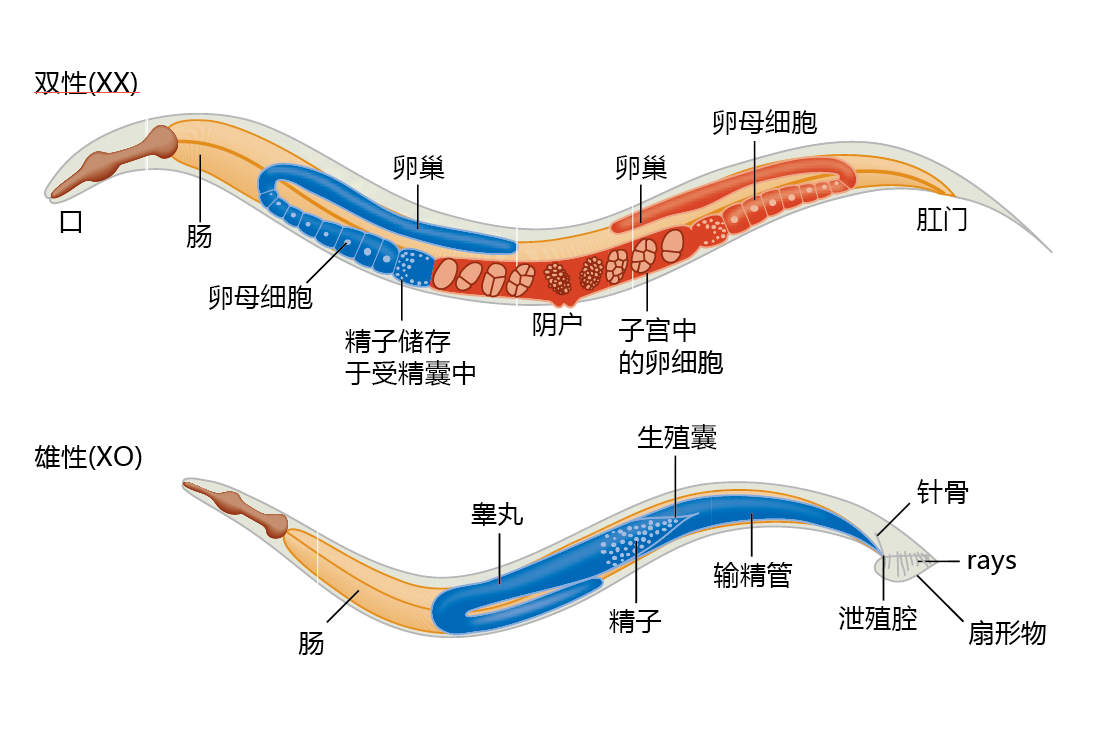
图5.35 秀丽隐杆线虫的两种性形态:雌雄同体和雄性。
成年雌雄同体的959个细胞都是有丝分裂后的;成年雄性由1031个有丝分裂后细胞组成。秀丽隐杆线虫完全处于有丝分裂后,并且具有显著的生殖后寿命,这使得秀丽隐杆虫成为研究多细胞生物寿命遗传调控的一个特殊模型。使用秀丽隐杆线虫的另一个重要优点是每个细胞的谱系和功能都是已知的。有趣的是,大约三分之一的细胞是神经细胞,这使得秀丽隐杆线虫在描述细胞调节和信号通路方面特别有用。
秀丽隐杆线虫的基因组大约有1亿对碱基,是酵母基因组的八倍,是果蝇基因组的四分之三。秀丽线虫有六条染色体,五对常染色体和一条性染色体。雌雄同体有两条性染色体(指定为XX)。雄性有一条X染色体(指定为XO)。雄性不能自己生育后代。然而,它们可以使雌雄同体杂交受精。基因组序列于1998年完成,约有20000个蛋白质编码基因。在本节中,我们学习dauer形成基因(daf-2),以及与daf-2相关的其他基因和时钟基因(clock gene)。
dauer基因的形成调节延长了寿命
秀丽隐杆线虫从卵到成虫的发育包括四个幼虫阶段,大约需要3-4天才能完成。成虫在成年后的头4天是生殖活跃的,在这4天的生殖期之后可能会再活10-15天。当秀丽隐杆线虫发现自己处于能够支持后代生存的环境中时,蠕虫会在正常的3至4天时间内,通过四个幼虫阶段进入成年期。然而,如果环境条件不适合繁殖,幼虫则在第3阶段停止发育,一种称作dauer的代谢活跃但繁殖沉默的幼虫形成(图5.36)。
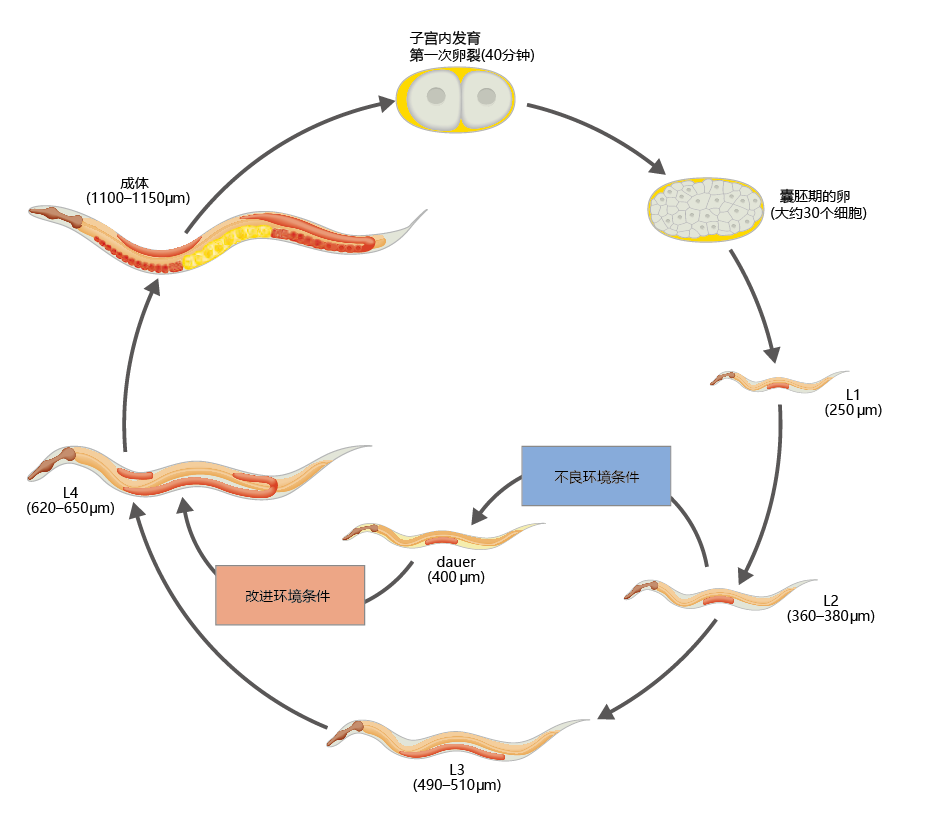
图5.36 秀丽隐杆线虫的发育阶段。卵在成年雌雄同体中受精,数小时后在大约30至40个细胞阶段。卵孵化后,蠕虫经过四个幼虫阶段(L1、L2、L3和L4),每个阶段都以蜕皮结束。成年后,每只秀丽隐杆线虫产生大约300个后代。它的寿命约为2周。如果第一阶段的环境条件不理想,那么第三阶段的生长就会停止,并形成一个dauer。秀丽线虫可以在这种停滞的发展状态下保持几个月。当环境条件改善时,dauer恢复发育成成虫。
在正常的生长周期中,性发育不成熟的dauer比第3阶段的正常个体小。在没有食物的情况下,以及在不适合繁殖的温度和土壤条件下,dauer可以存活数月。它通过降低代谢率、限制蛋白质合成和依靠储存的脂肪生存来完成这一奇迹过程。dauer的解剖学变化增加了它对环境压力的抵抗,从而提高了它的生存机会。这些变化包括(1)增加角质层的厚度;(2) 在某些情况下,闭合口腔;(3)增加内源性抗氧化剂的浓度。这些形态变化是如此有效,以至于秀丽隐杆线虫能够在洗涤剂、辐射和大量其他有害物质的伤害下存活数小时。尽管如此,当环境条件改善时,dauer可以迅速恢复发育。遗传和生物化学机制允许dauer在环境中放置食物后1小时内重新进入幼虫阶段;幼虫4期的蜕皮发生在喂食后的8小时内。
遗传途径调控dauer的形成
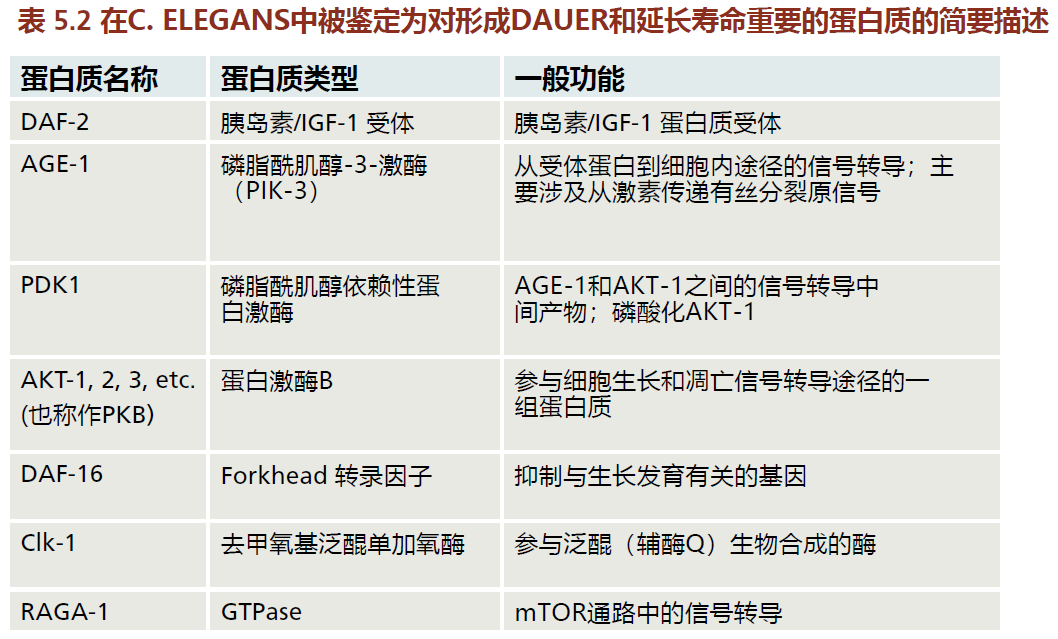
如表5.2所示,科学家已鉴定出几种在秀丽隐杆线虫dauer的形成中具有重要作用的蛋白质。参与dauer形成的遗传途径和dauer在极端压力下生存的能力为延长寿命的机制提供了一些重要的信息。在秀丽隐杆线虫突变体的遗传繁殖过程中,科学家已经鉴定出两个关键基因,这两个基因的信号通路是相同的,分别是正常生长和繁殖所需的age-1(因其衰老表现型而得名)和dauer形成基因daf-2,daf-2是正常生长和繁殖必须的。没有这些基因,秀丽隐杆线虫会形成dauers。age-1基因的克隆表明其蛋白产物(AGE-1)是高度保守的磷脂酰肌醇-3-激酶(PIK-3)家族的成员。这些蛋白质是膜受体之间的重要细胞内中间体,它们是启动信号和细胞内活动所必须的。daf-2基因的克隆表明,它编码一种跨膜受体蛋白(DAF-2),与其他物种中发现的胰岛素/胰岛素样生长因子(IGF-1)受体具有同源性。一个高度保守的基因daf-2,就像它的遗传同源基因一样,对正常的生长和繁殖很重要。
在环境充足的时期,传递给蠕虫的信号导致DAF-2结合蛋白的表达(图5.37)。DAF-2蛋白与DAF-2受体的结合导致AGE-1蛋白从细胞质内部迁移到细胞膜。这一发现提供了一个重要的线索,即dauer的形成是由与内分泌信号相关的基因的抑制所调节的。也就是说,磷脂酰肌醇-3-激酶在很大程度上响应于激素而被激活。
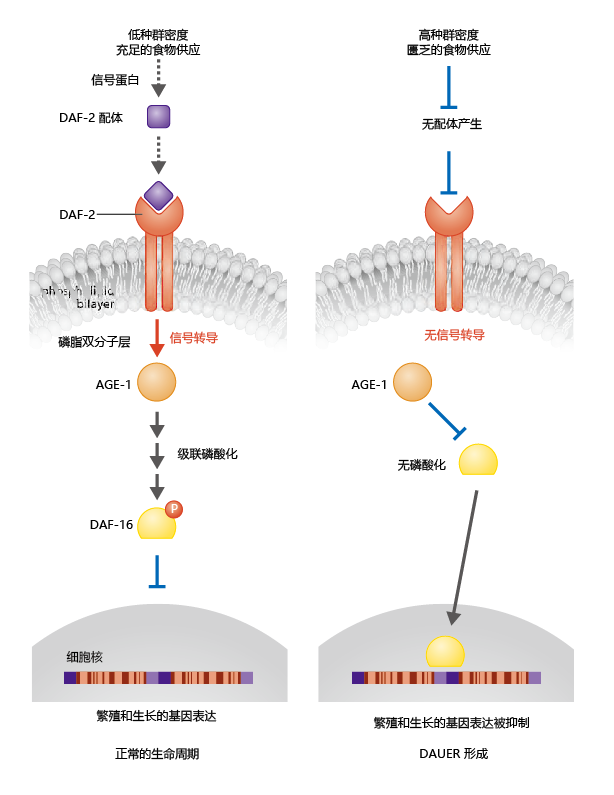
图5.37 秀丽隐杆线虫成虫繁殖和dauer形成中基因表达的调节。最佳环境条件刺激胰岛素样配体与DAF-2受体的结合(左)。信号通过膜传播,并将细胞内AGE-1(一种磷脂酰肌醇-3-激酶)吸引到膜上,并且在膜上激酶启动磷酸化级联反应。该级联反应以DAF-16的磷酸化结束,DAF-16是一类参与调节生殖的蛋白质。DAF-16的磷酸化阻止了它进入细胞核,在没有DAF-16的情况下,参与生殖和生长的基因得以表达。恶劣的环境条件阻止了DAF-16的磷酸化,DAF-16进入细胞核并抑制了与生殖有关的的基因(右);进而引起了dauer的形成。
当研究人员发现DAF-16的磷酸化是正常生长和繁殖所必需的时,我们对秀丽隐杆线虫dauer形成及其与寿命延长的关系的理解取得了下一个重大进展。如图5.37所示,DAF-16的磷酸化阻止它从细胞质迁移到细胞核。因此,DAF-16不能对那些生长和繁殖的基因发挥其抑制作用;秀丽线虫在四个幼虫阶段正常发育至成年,其正常寿命为10-15天。如果环境条件不足以支持生殖,则不会合成DAF-2结合蛋白,也不会通过DAF-2受体将信号转导至AGE-1。如果DAF-16蛋白没有磷酸化,那么它就会进入细胞核,在细胞核中它抑制了调节生长和繁殖的基因。如果DAF-16对基因的抑制发生在幼虫第3阶段,则形成dauer。
daf-16基因的克隆表明,daf-16蛋白是叉头盒(forkhead box)转录因子家族(FOXO)的成员。这种特殊的转录因子抑制秀丽隐杆线虫在低食物条件下生长和繁殖的基因。FOXO是一组高度保守的蛋白质,参与许多不同物种的繁殖,包括几种哺乳动物。
daf-2弱突变延长寿命
调节dauer形成的关键基因中的弱突变使得成年个体寿命延长。弱突变是指基因的改变,基因表达减少而不是完全消除。当在daf-2中诱导弱突变时,DAF-16的磷酸化减少但不被阻止。与野生型相比,第三阶段幼虫的新陈代谢略有降低,不会形成dauer。相反,DAF-16磷酸化的减少使秀丽隐杆线虫能够在幼虫发育成成虫的四个阶段正常发育。虽然向成年期的进展可能是正常的,但突变的影响是非同寻常的:daf-2弱突变的秀丽隐杆线虫比野生型寿命长50%-300%。
秀丽隐杆线虫延长寿命突变体的表现和行为在很大程度上与正常寿命的成虫一样。成年蠕虫仍能生育,雌雄同体只产生少量后代,尽管对突变的一些轻微修改可能导致不育,但这表明繁殖和延长寿命之间存在密切关系。与dauer不同,含有daf-2弱突变的成虫蠕虫可以正常进食,并对环境变化(包括温度和接触)做出强烈反应。
daf-2基因将长寿与神经内分泌控制联系起来
daf-2基因的克隆和DAF-2蛋白作为胰岛素/IGF-1样受体的鉴定表明,引起成年蠕虫寿命延长的基因调控涉及神经内分泌控制。当发现信号转导途径涉及PIK-3(AGE-1)时,daf-2神经内分泌调节的进一步证据就出现了,这是一种磷酸化机制,通常在细胞周期的激素调节中发现。进一步的实验证实,如果在神经细胞和肌肉细胞都有daf-2突变,那么只有神经组织的daf-2突变才能引起环境感知和信号转导,进而引起寿命的延长。当突变仅限于肌肉细胞时,寿命没有增加。然而,将突变局限于神经内分泌细胞时可以延长寿命。也就是说,神经内分泌机制似乎调节秀丽隐杆线虫的dauer形成和延长寿命。
DAF-2蛋白参与衰老的神经内分泌机制的进一步证据来自实验室研究,研究表明,mTOR2途径对胰岛素/IGF-1样受体的信号作出反应(图5.38)。虽然对mTOR2的了解远少于mTOR1,但我们确实知道,功能正常的mTOR2通路对细胞生存至关重要。大多数研究表明,mTOR2有助于维持细胞骨架(细胞结构)并抵御细胞压力。因此,毫不奇怪的是,如果剔除mTOR2途径的各种成分的基因将会导致秀丽隐杆线虫的寿命缩短。然而,mTOR2途径中的弱功能缺失突变(见上文)似乎会延长寿命。这一有趣的悖论尚未得到解释,但一些研究表明,抑制mTOR2通路的一部分可能会抑制先前描述的DAF-2/DAF-16通路。衰老的神经内分泌机制将在接下来的两节中详细讨论。
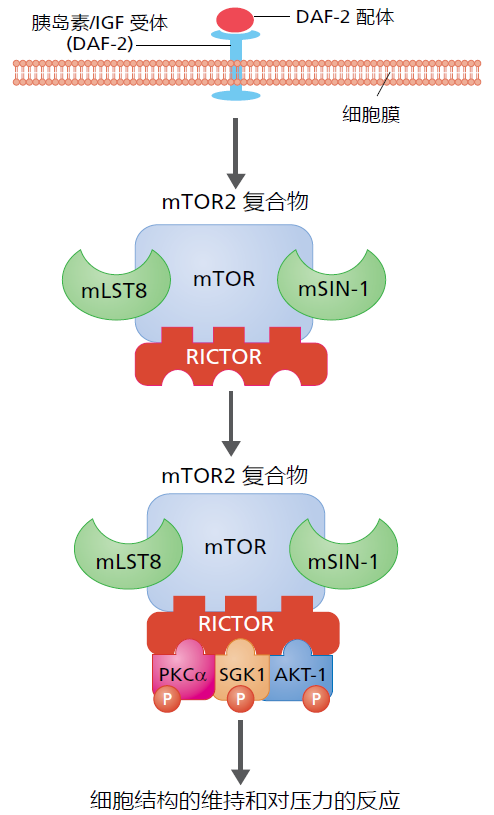
图5.38 mTOR2通路的简化图示。DAF-2配体与蠕虫中的DAF-2受体(胰岛素/IGF受体)的结合引起mTOR2复合物的形成。RICTOR是一种支架蛋白,为mTOR2复合物与其靶蛋白的结合提供空间。附着于mTOR蛋白上的mLST8和mSIN-1有助于靶蛋白PKCa、SGK-1和AKT-1的磷酸化。PKCa、SGK-1和AKT-1的磷酸化会激活对细胞生存至关重要的其他信号通路。
线粒体蛋白可能与延长寿命和新陈代谢有关
秀丽隐杆线虫的能量代谢和寿命延长之间的直接联系已被确定为另一种基因调节途径,它们是线粒体中发现了时钟基因(clk-1、clk-2、clk-3和gro-1)。时钟基因是一组高度多效性的基因,之所以如此命名,是因为它们调节着几种线粒体功能的定时和同步。clk-1的功能缺失突变会影响好几个系统,并导致许多功能的“减缓”,包括幼虫发育、卵子生产和幼崽大小。clk-1突变体功能的减缓也产生了一种比野生型秀丽隐杆线虫寿命长15%–30%的稍小的蠕虫。由于其多效性,最有可能与长寿有关的clk-1基因途径尚未被确定。
最普遍接受的关于时钟基因的理论是将新陈代谢速度的减缓与长寿联系起来。Clk-1蛋白被鉴定为脱甲氧基泛醌单加氧酶(DMO),它是一种泛醌生物合成所需的酶,也称为辅酶Q(CoQ)。辅酶Q在线粒体内膜中高浓度存在,在那里它充当黄素蛋白和细胞色素b之间的电子转移蛋白。clk-1功能缺失突变会导致辅酶Q减缓了电子通过ETS的转移,但不会停止。反过来,能量储备下降,生理机能必须减慢。
从clk-1调节DMO基因表达的途径尚未描述,寿命延长的机制也未描述。然而,以线粒体中CoQ的功能为中心的两个理论已经建立。一种理论认为,CoQ水平的降低会减少ETS活性,进而减少以氧为中心的自由基的产生。自由基的减少会减少细胞损伤,并可能延长寿命。然而,这一解释并不能解释clk-1基因在几种不同系统的减缓中的多效性效应。第二种理论预测,细胞核中的调节过程“感知”细胞的慢性低能量状态,即低ATP导致参与各种生理功能的蛋白质(一种能量恒温器)的基因表达减少。该理论解释了clk-1基因的多效性,但未能解释延长的寿命。与许多延长秀丽隐杆线虫寿命的基因一样,我们需要对时钟基因如何延长寿命的调控基因表达的途径要有一个更加精细的特征描述。