
5.6 果蝇寿命的遗传调控
酿酒酵母和秀丽隐杆线虫等简单生物的单个基因突变表明,长寿可能具有高度调控的遗传元件。这些简单生物的使用得益于这样一个事实,即它们的大多数基因都具有单一的功能,并且是响应于单一的生化途径或环境刺激而表达的。也就是说,与更复杂的生命形式相比,引起寿命改变的基因调控途径可以在这些简单的生物体中相当容易地分离出来。当我们在进化阶梯上前进时,解剖学和生理学的复杂性水平是基因表达调控水平提高的结果。复杂生物中的基因往往是高度多效性的,它们影响多种生理功能,任何一种基因的表达都可以通过多种途径调节。因此,在复杂的生物体中,分离和描述长寿的遗传调控途径往往比在简单的生物体(如酿酒酵母或秀丽隐杆线虫)中更困难。
对诸如黑腹果蝇等复杂生物寿命的遗传调控的描述,在很大程度上局限于对基因选择品系的寿命分析。这些结果清楚地表明,许多不同基因位点的突变可以延长寿命(表5.3)。这些结果虽然支持长寿的遗传调控的可能性,但不能最终证明编码特定蛋白质的特定基因直接引起寿命延长。鉴于我们对基因是如何被调控的新理解(参见前面的“基因表达调控”一节),引起长寿改变的突变基因可能只是更复杂途径的一部分。因此,为了尽量减少基因对寿命影响的讨论中不必要的猜测和不确定性,我们对复杂生物体长寿的遗传调控的探索主要集中在从酵母到人类的高度保守的途径上。这些途径与神经激素信号传导有关,类似于秀丽隐杆线虫的实验。我们首先简要探讨延长寿命的果蝇突变体,为更深入的分析奠定基础。然后我们讨论了三种途径,在果蝇中,它们证明了神经激素信号传导与寿命之间的联系。

果蝇在基因研究方面有着悠久的历史
我们对经典遗传学和分子遗传学的了解大多来自对黑腹果蝇的研究。100多年前就有报道称,首次成功培育出一种具有特定特性的果蝇。托马斯·亨特·摩根(Thomas Hunt Morgan,1866–1945)利用果蝇发现X染色体中含有控制果蝇红眼和白眼的基因,他于1933年获得了诺贝尔医学奖,这是遗传学家首次获得该奖项。调节身体部位位置的Hox基因(见方框5.1)在所有多细胞生物中几乎是相同的,这一发现在很大程度上来自对果蝇的研究。
果蝇基因组包含约1.25亿个碱基对,其中约20%构成15000–17000个基因。DNA包含在四对同源染色体之中,其中三对常染色体,一对性染色体。果蝇的生命周期由六个阶段组成:胚胎发生、三个幼虫阶段、蛹阶段和成虫阶段(图5.39)。在理想的温度条件(25°C)和充足的食物下,从卵到成虫的发育时间大约需要9天,但如果温度和食物低于最佳值,则可能需要更长的时间。雄性比雌性小。雌性在羽化后12小时便可受精,并可在15-25天内保持生育能力。野生型果蝇的平均寿命为40-50天。
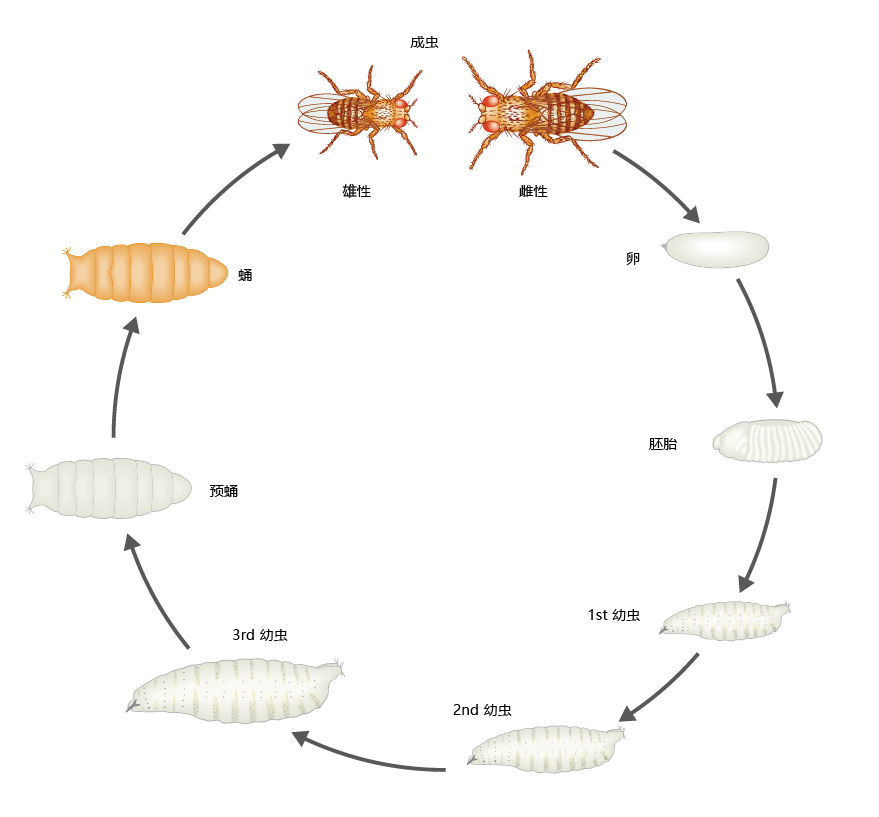
图5.39 果蝇生命周期的阶段。
延长寿命的基因与增加的抗压能力有关
表5.3列出了一些被鉴定为延长果蝇寿命的基因。这些基因中的许多都有一个共同的功能,那就是抗压力。例如,hsp70编码的伴侣蛋白(热休克蛋白)属于一组标记错误折叠或受损蛋白通过泛素途径降解的蛋白质(参见本章前面的“蛋白质可以在翻译后被修饰或降解”)。在果蝇和小鼠中完成了大量实验表明,hsp70的过度表达可以延长寿命。另一组参与抗压功能的蛋白质是内源性抗氧化剂,这种蛋白质能降解以氧为中心的自由基,并多次被证明对果蝇的衰老速度和寿命有重大影响。超氧化物歧化酶是将超氧化物自由基还原为水的一种酶,其过度表达可以(但并非总是)延长了果蝇的寿命。很明显,热休克蛋白和内源性抗氧化剂与果蝇延长寿命有关,但它们的表达是对其他细胞内信号的反应。也就是说,这两种蛋白质基因上游的一些基因一定是长寿的原因。这些基因有待鉴定。
随着Methuselah(mth)基因的发现和克隆,抗压力基因与长寿之间的联系变得更加清晰。(据报道,Methuselah是一位圣经人物,活了969年。)仔细筛选果蝇突变体,发现了一种耐热的长寿菌株。转基因mth果蝇的寿命明显长于野生型(图5.40),在抵抗百草枯(一种诱导氧中心自由基过度产生的化学物质)、饥饿和高温的胁迫方面更有效(图5.41)。mth基因的克隆显示出与G蛋白偶联受体的显著同源性。回想一下,在酵母中,G偶联受体已被鉴定为具有延长寿命的作用。当G偶联受体与特异性配体的结合延长了转基因mth果蝇突变体的寿命时,G偶联受体在长寿中的中心作用被证实。
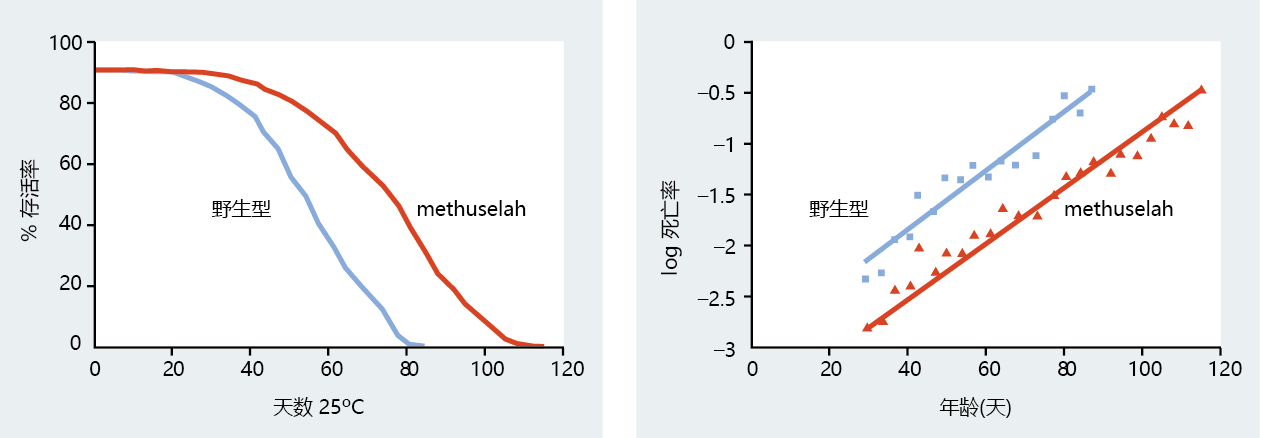
图5.40 野生型和mth转基因果蝇的存活曲线和死亡率对数。注意,mth转基因品系的死亡率对数向右偏移,表明衰老速度已经减慢。(摘自Lin YJ等人,1998年。科学282:943–946。经AAAS许可。)
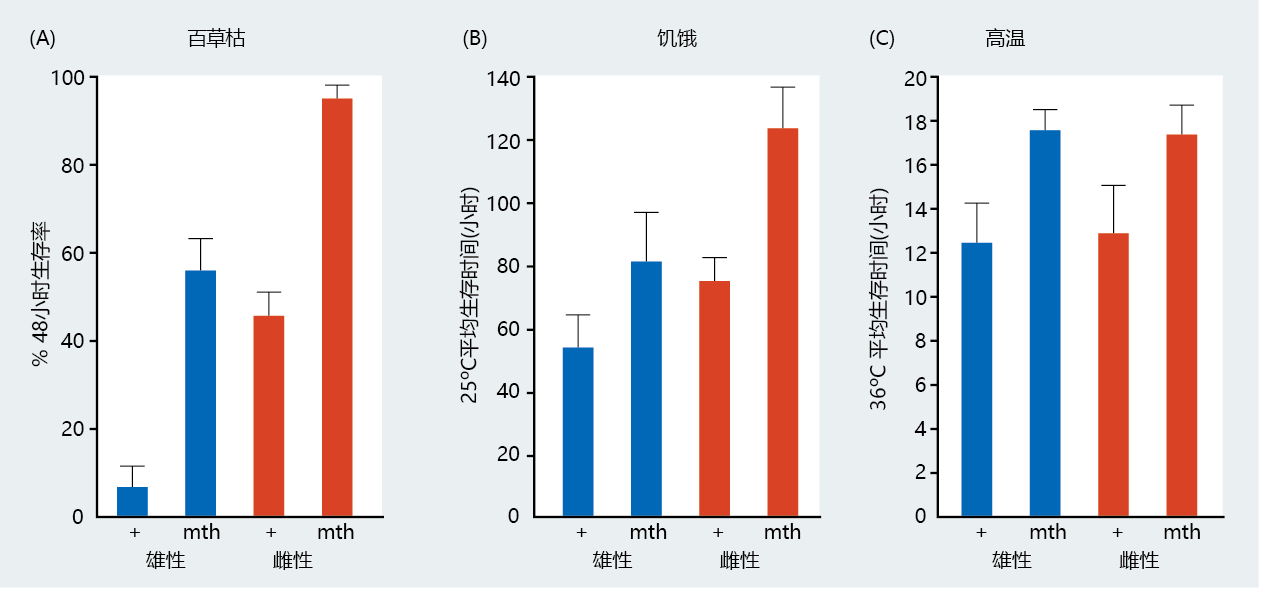
图5.41 果蝇对压力的反应存活。这些图表显示了雄性和雌性野生型(+)和mth转基因果蝇对(A)百草枯、(B)饥饿和(C)高温的反应。在所有治疗中,携带mth突变的雄性和雌性苍蝇比对照野生型苍蝇存活时间更长。(摘自Lin YJ等人,1998年。科学282:943–946。经AAAS许可。)
控制果蝇生长的基因也能延长寿命
酿酒酵母和秀丽隐杆线虫的长寿似乎与环境条件密切相关,环境条件反过来影响繁殖。对动物造成压力的环境条件(例如食物缺乏)会抑制基因表达,进而导致生长和繁殖停止,基因表达的抑制直到环境对后代的生存更有利才解除。抑制酿酒酵母和秀丽隐杆线虫生长和繁殖的信号通路也延长了寿命。从秀丽隐杆线虫的实验中发现,启动dauer形成和延长寿命的信号来自神经内分泌。和许多昆虫一样,果蝇也开发出了一种在环境条件恶劣时延缓繁殖的策略。这种被称为滞育(diapause)的策略似乎受到神经激素信号的严重影响,其特点是生殖沉默、能量代谢降低和抗压。生物衰老学家利用果蝇的滞育来评估影响寿命的基因。
一种被称为chico(西班牙语“小男孩”)的果蝇突变株提供了进一步的证据,该证据表明,比秀丽隐杆线虫更复杂的中,与生长和繁殖相关的胰岛素相关基因调控途径可能在衰老速度中起重要作用。chico果蝇的大小大约是野生型果蝇的一半;大小差异是这种突变体细胞越来越少的直接结果(图5.42)。chico突变体包含一个功能缺失突变(敲除),该突变降低了胰岛素受体底物蛋白CHICO的表达。chico基因与秀丽隐杆线虫的daf-2突变体高度同源,并且已表明chico的表达减少会延长果蝇的平均寿命(图5.43)。此外,CHICO蛋白通过类似于秀丽隐杆虫中描述的途径刺激生长:抑制叉头转录因子dFOXO。

图5.42 雌性chico突变体(左)和野生型(右)成年果蝇。(摘自Piper MD等人,2008年。《内科实习杂志》263:179–191。经Wiley许可。)
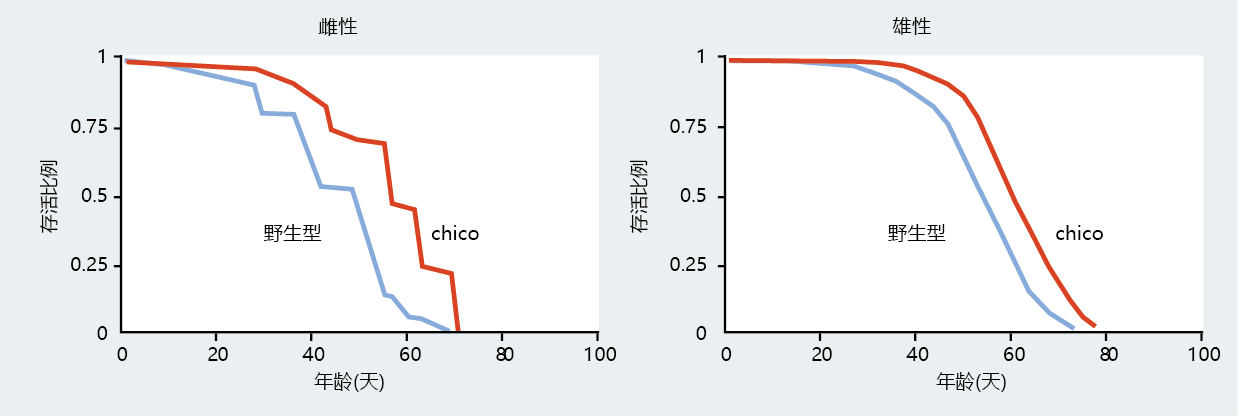
图5.43 具有和不具有chico基因的雌蝇和雄蝇的寿命。chico基因敲除的存活率仅限于平均寿命(Increased survival of the chico knockout was limited to mean life span. )。这可能表明,与直接增加寿命相比,chico在减缓衰老速度方面发挥了更大的作用。(摘自Clancy DJ等人,2001年。科学292:104-106。经AAAS许可。)
涉及胰岛素途径的进化保守机制可能参与寿命调节的可能性得到了进一步的支持,发现胰岛素途径受体InR的功能缺失突变也增加了寿命。这种突变导致激酶样活性显著降低。这种功能丧失突变体的一个独特特征是,与野生型果蝇相比,突变体的卵子产量显著减少,这一发现将这种突变与滞育联系起来(图5.44)。当发现保幼激素(JH:一种参与幼虫发育的激素)恢复成年果蝇InR功能并降低寿命时,生殖滞育与促进长寿之间的关联就得到了进一步的证实。此外,对果蝇的一些研究表明,mTOR途径的功能丧失抑制了胰岛素/胰岛素样生长因子途径,引起寿命延长。mTOR和胰岛素/胰岛素样生长因子途径之间的关系是复杂的,需要进一步研究。
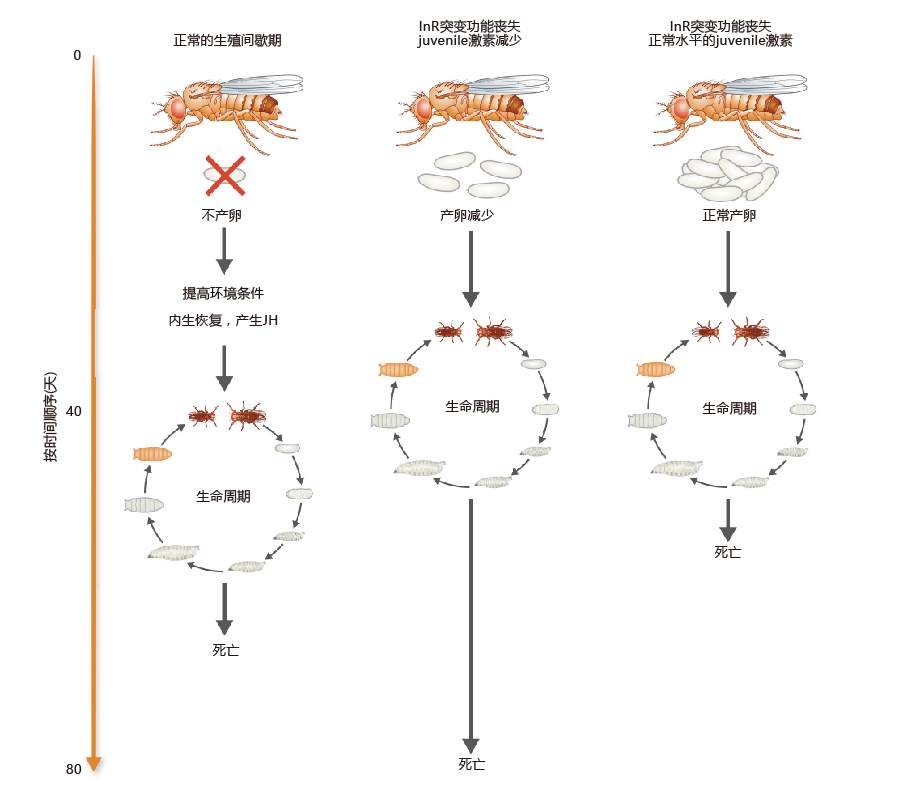
图5.44 生殖滞育、保幼激素(JH)和寿命之间的关系。处于繁殖滞育期的果蝇不产卵。当环境条件改善时,产卵恢复,相对于野生型苍蝇,繁殖滞育期的果蝇寿命延长。胰岛素样受体(InR)功能缺失突变降低了卵子产量并延长了寿命(中)。用JH处理InRmutated果蝇可以恢复正常的产卵量(右),但这种处理会缩短寿命。