
1.4 衰老生物学家们怎么研究衰老:比较衰老生物学
Although research on laboratory species has provided, and continues to provide, significant insight into the basic biological mechanism of aging, the short life spans of these species make them less effective as models for investigating the mechanisms that underlie the exceptional longevity observed in humans. Some biogerontologists study longevity by observing wild animals that have long life spans, a subfield of biogerontology called comparative biogerontology. Comparative biogerontology identifies wild species that show resistance to aging and thus have extended longevity in 环境s that are otherwise conducive to short life spans. The long-lived species can then be bred in captivity, where possible genetic and biochemical mechanisms underlying their extended longevity can be evaluated. Identification of the mechanisms that have allowed the 进化 of resistance to early death and thus lengthened life spans provides clues as to how extended longevity evolved in humans.
Our focus here is on a general overview of comparative biogerontology, describing a few relationships suggested to extend longevity in wild animals. We also describe a few specific animals with extended longevity and the 进化ary adaptations that have provided these animals with long life spans in the wild. The 进化ary, genetic, and biochemical mechanisms accounting for extended longevity are covered thoroughly in Chapters 3, 4, and 5.
物种的身体尺寸与最大寿命是相关联的
A field mouse‘s maximum life span is much shorter than a rabbit‘s; a rabbit‘s much shorter than an elephant‘s. The casual observation that large mammals live longer than smaller ones was first noted in the scientific literature more than a hundred years ago and was confirmed in more recent studies (Figure 1.13) . The relationship between body size—here meaning overall dimensions, independent of underweight or overweight—and longevity also holds when warm-blooded animals are separated into their taxonomic groups, such as primates, ungulates, carnivores, rodents, and so on, and within the avian class. Humans, interestingly, do not fit within this pattern, having the longest life span of any mammal, yet clearly not the largest mammal. Nonhuman primates also fall outside the range in the body size–versus–life span curve established for other mammals. The unique position of the primates, including Homo sapiens, within the body size–life span relationship most likely reflects factors involving intelligence; this is discussed more thoroughly in Chapter 7.
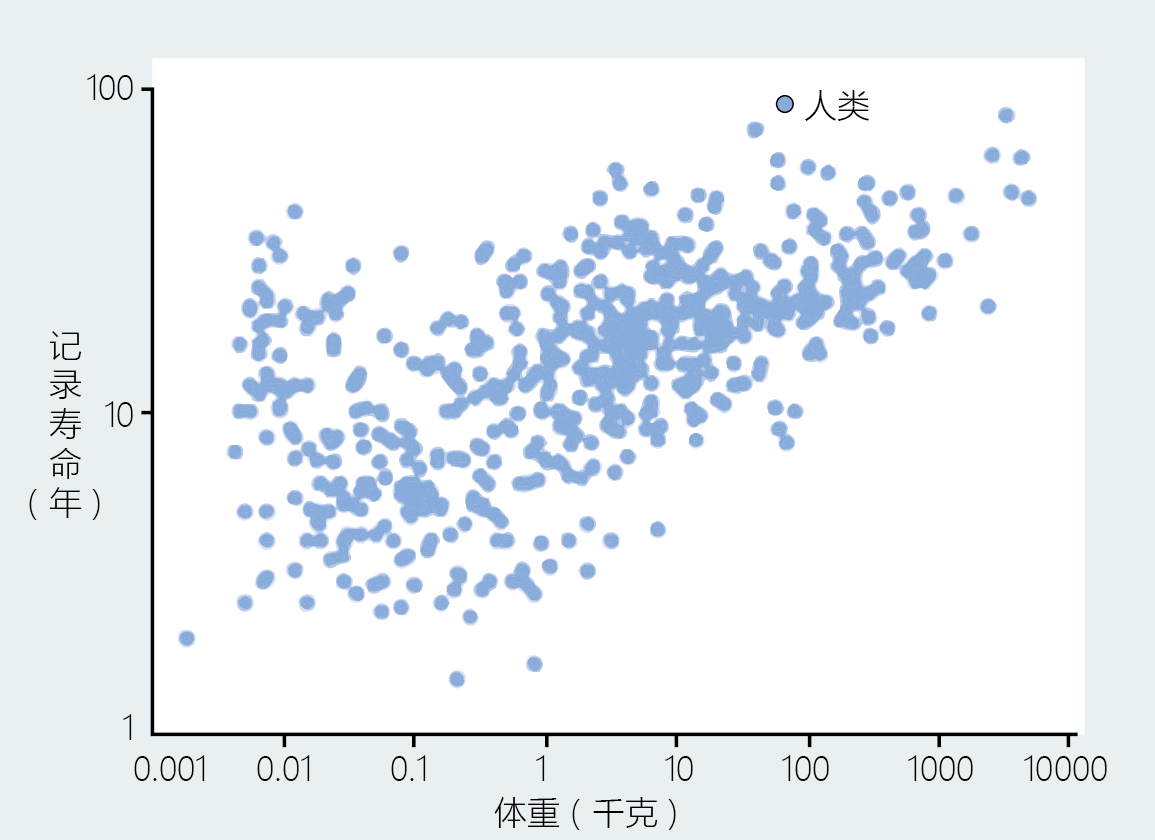
Figure 1.13 Relationship between life span and body mass in 605 mammalian species. Note that humans have the longest life span among the mammals, despite having a smaller body mass than some other mammals. (Adapted from S. Austad, in The Comparative Biology of Aging [N.S. Wolf, ed.], New York: Springer Science, 2006.)
Humans' large brains, combined with their exceptional longevity among mammals, suggested to many scientists that brain size might be the factor that accounts for the body size–life span relationship: larger animals tend to have larger and more complex brains. Such a hypothesis would seem reasonable, given that larger brains tend to provide more intelligence and greater regulation of the physiological functions that help to maintain homeostasis (the ability to maintain internal stability). Intelligence would help animals escape predators and enhance their success at finding food. Superior control of homeostasis would allow animals to exist over a larger range of temperatures and to survive in many different types of 环境al conditions. These animals would then have a bigger range over which to find food and thus increase their chances of survival.
Although the brain–weight hypothesis has innate appeal and has been supported in some investigations, the vast majority of studies do not find that brain weight significantly accounts for the body size –life span relationship. Indeed, the sizes of organs such as the liver, spleen, and heart are better predictors of longevity in most mammals than is brain size. Since the sizes of most internal organs other than the brain are determined, in large part, by the overall size of the body, it is not surprising that the size of internal organs would be as good a predictor of life span as body size.
The physiological complexity of the warm-blooded animals (also known as homeothermic and endothermic animals) has led biogerontologists to suggest that some factor (or factors) other than the simple measure of size may more closely reflect the relationship between longevity and body size. For example, scientists exploring the relationship between body size and longevity in the first half of the twentieth century noted that smaller mammals have significantly faster rates of metabolism than larger mammals, as measured by daily energy expenditure per total body mass. This led to a general theory suggesting that the greater the rate of energy expenditure, the shorter the life span—known scientifically as the “rate-of-living theory” and more popularly as “live fast, die young.”
The rate-of-living theory as a general explanation for the body size –longevity relationship has not held up under more extensive and rigorous scientific experimentation, although it remains popular in the nonscientific literature. Animals of the avian class have metabolic rates twice those of mammals with a similar body mass (Figure 1.14) . However, birds live two to three times longer than their body size–matched mammalian counterparts. Marsupials, nonplacental mammals (for example, kangaroos, opossums), have shorter life spans and slower metabolic rates than comparably sized eutherians, the placental mammals.
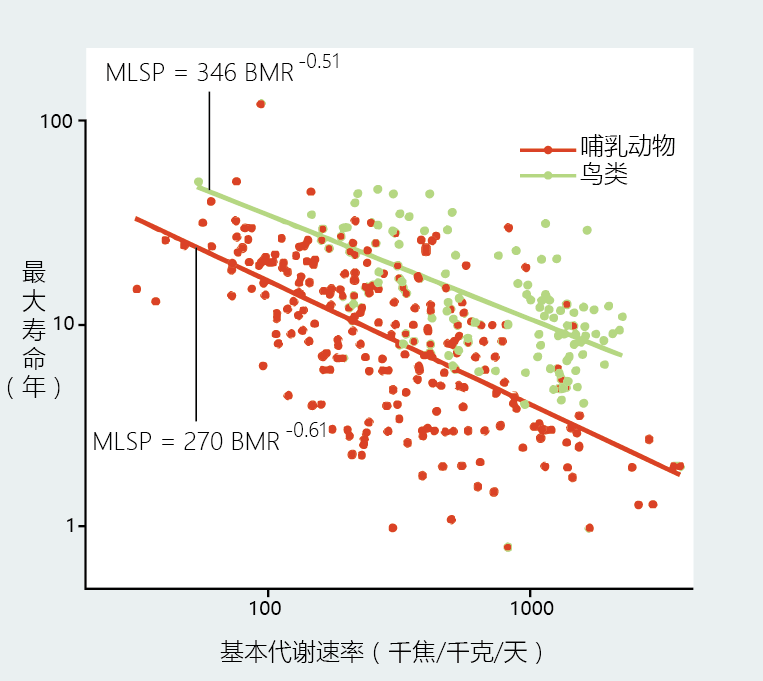
Figure 1.14 Relationship between basal metabolic rate (BMR) and maximum life span potential (MLSP) in birds and mammals. Birds have a longer life span and greater metabolic rate than mammals of similar body size. (From A.J. Hulbert, R. Pamplona, R. Buffenstein, and W.A. Buttemer, Physiol. Rev. 87:1175–1213, 2006. With permission from the American Psychological Society.)
Reduced vulnerability to extrinsic dangers explains extended 寿命
Although observation and statistical analyses suggest that body size may be related to extended longevity in the wild, sufficient numbers of exceptions have been found among warm-blooded animals to suggest that this relationship is casual, not causal. The causal factor (or factors) that allows for extended longevity in wild animals, although not yet fully described, most likely reflects species‘ ability to adapt to their environment. Large animals may have extended longevity simply because their size and strength make them better at defending themselves against predators. Large animals also tend to have much larger foraging areas than small animals. This would reduce population density and decrease competition for food. Over 进化ary time, the ability to defend against predation and to decrease the risk of starvation would enhance survival and lead to the 进化 of extended longevity. The underlying mechanism for the 进化 of extended longevity is related to delayed reproduction, as discussed in detail in Chapter 3.
An ability to escape predation and to forage over a larger area as the mechanism for the 进化 of long life spans in the wild can also be illustrated by animals that fly. Nonflying birds or weak-flying birds (chickens, for example) have significantly shorter life spans than strongflying birds. Flight is an extremely effective way to escape predators and a more efficient mechanism for traveling long distances than the use of legs—thus providing a greater foraging area. Moreover, bats are the longest-lived mammals for their size.
Nonflying mammals and cold-blooded animals with expanded longevity share an ability to protect themselves from predation. The quills of the porcupine are most likely the reason that these mammals are thought to be the second longest lived (for body size), after the bats. Some species of turtles are known to live more than 150 years.
A highly organized social structure also extends 寿命 in the wild
Safety in numbers is another aspect of the 进化 of extended life span in wild species. Social animals such as primates and herd animals consistently have longer life spans for their body size than nonsocial animals. An excellent example of societal influence on longevity occurs in the naked mole rat of Equatorial Africa (Figure 1.15) . The naked mole rat is the size of a mouse and lives out its life completely underground. That is, naked mole rats are never exposed to the dangers of the terrestrial world, and they live within large colonies. As a result, these animals live for 20 –30 years, two to three times as long as other mammals of similar size.

Figure 1.15 The naked mole rat (Heterocephalus glaber). Naked mole rats spend their entire lives underground, reducing their exposure to the harsh 环境. This may be a factor in their relatively high longevity for their body size. (From R. Buffenstein J. Gerontol. A. Biol. Sci. Med. Sci. 60:1369-1377, 2005. With permission from Oxford University Press.)
Social insects, such as termites, ants, wasp, and bees, are another group of animals that have developed extended longevity. The social aspects of these insects involve a division of labor based on reproductive function, a type of social organization known as eusociality. The reproductive function also determines the life span of the insects. For example, each hive of honeybees has only one reproductively active queen, which can live 5–7 years. The queen is genetically identical to the thousands of functionally sterile female workers that attend to the larvae and pupae and live for just a few months. Some of the female workers will transition into foragers, the bees that collect pollen and make honey. These female foragers have life spans of less than 30 days. Finally, the male drones, whose only job is to mate with the queen, live through only one seasonal cycle. Thus, one colony of bees contains genetically identical females with three different phenotypes and life spans and males whose life span is directly related to reproduction (Figure 1.16) . Moreover, the castes into which the females are assigned appear to be controlled by nutritional status during development. The best-nourished larvae become queens. This unique feature of the eusocial insects provides biogerontologists with a method to easily manipulate life span and evaluate the interaction between nutrition and longevity.

Figure 1.16 Morphology of honeybees. The morphology and life spans of honeybees appear to be related to differences in nutrition during the larval phase. (A) A queen bee (the large bee in the middle) and workers (the smaller bees surrounding the queen). (B) A male drone. (A, courtesy of angelshot/Shutterstock; B, courtesy of alle/Shutterstock.)
A few aquatic animals have extreme 寿命
Sponges, jellyfish, sea anemones, clams, and some fish, as noted earlier in the chapter, are thought to have extreme longevity, although precise age estimates are difficult to make, as most of these species have not been maintained in the laboratory. White sturgeon (Acipenser transmontanus), a freshwater fish found on the west coast of North America, has been estimated to live upward of 200 years. Unpublished accounts of sea anemones living more than 150 years in a fish tank are often cited. Only recently has confirmation of extreme longevity in a clam, Arctica islandica, been documented. Carbon-dating of the shell places one clam at 400 years of age and many others at 100 years (Figure 1.17) .

Figure 1.17 Extreme longevity in aquatic animals. (A) The maximum life span of the white sturgeon (Acipenser transmontanus) is unknown but is estimated to be close to 200 years. (B) A clam of this Arctica islandica species was carbon-dated as 400 years of age. (A, courtesy of Shutterstock; B, courtesy of Z. Ungvari, Z. Ungvari et al., J. Gerontol. A Biol. Sci. Med. Sci. 66:741 –750, 2011. With permission from Oxford University Press.)
The biochemical and genetic mechanisms underlying the extreme longevity in these aquatic species have yet to be determined. However, this extreme longevity seems to be associated with continual growth. As you will learn in Chapter 3, growth and development are associated with many biological functions that enhance survival. The long-lived clam shows a heightened resistance to cellular damage, the basic mechanism underlying aging. In sponges and jellyfish, a unique mechanism has evolved that may account for their extreme longevity. The cells of sponges and jellyfish can transition between somatic cells (cells not involved in sexual reproduction) and germ cells (reproductive cells, produced by the sex organs or tissues of multicellular organisms, that transmit hereditary information). You will learn in Chapter 4 that germ cells are extremely efficient at protecting themselves from damage and are thought to have infinite life spans.
ESSENTIAL CONCEPTS
- Biogerontology explores the biological processes that occur inside living things as they age and integrates research from many different scientific fields.
- Biogerontology emerged as an independent field of study in the 1940s, after scientific and technological advances led to a significant increase in the average human life span.
- Aging research is unlike any other health-related research in that aging and death have no cure.
- Aging reflects the result of a lifetime of interactions with our 环境, and no two humans have the same interactions with their 环境.
- The differences in the rate of aging within and between species make research in aging and longevity challenging and have presented an obstacle to a precise definition of aging.
- There are many different definitions of aging, each having a particular place in the study of aging. For this textbook, aging is defined as random change in the structure and function of molecules, cells, and organisms caused by the passage of time and one‘s interaction with the 环境. Aging increases the probability of death.
- Development, maturity, and senescence are specific eventrelated points, or stages, within the life span that describe distinct periods of 生物衰老 that can be used to make comparisons of changes across the life span.
- Aging and disease are separate entities. A disease is a process within the animal or plant that impairs normal function. Aging occurs within the normal bounds of biology.
- Because ethical and practical considerations limit the type of research that can be done in humans, biogerontologists use a variety of laboratory organisms, from single-cell organisms, invertebrates including insects, a range of mammals and fish, birds, nonhuman primates, to understand the basic nature of human aging. However, no single animal or plant model can be seen as a “perfect” system for studying 生物衰老.
- Investigations of exceptional longevity of some wild animals are used to suggest 进化ary mechanisms that may apply to human longevity. The study of aging in the wild is called comparative biogerontology.
本章结束。