
4.2 细胞周期的调节
对于生物来说,没有什么比DNA的准确复制和正确的细胞分裂更重要的了。如果出问题会引起细胞功能异常,或者是疾病,例如癌症。真核细胞进化出一套复杂的系统调节细胞分裂,保证DNA的准确复制。这些调节主要是各种各样的“检查点”,这些“检查点”判断是否进行下一步。如果调节系统功能正常并探测到DNA错误,染色体压缩以及细胞分裂就会停止,直到DNA错误被修复。这些检查点调节使得DNA复制暂停的现象通常叫做分子闸(molecular brakes)。
本部分我们简要讨论细胞周期调节的分子机制,这个过程通常被称作细胞周期控制系统。我们按照细胞周期的顺序逐个讨论调节方式,从细胞外的触发细胞周期的信号,到胞质分裂期子细胞的分离。
4.2.1 S-细胞周期蛋白类和细胞周期蛋白依赖的激酶触发DNA复制
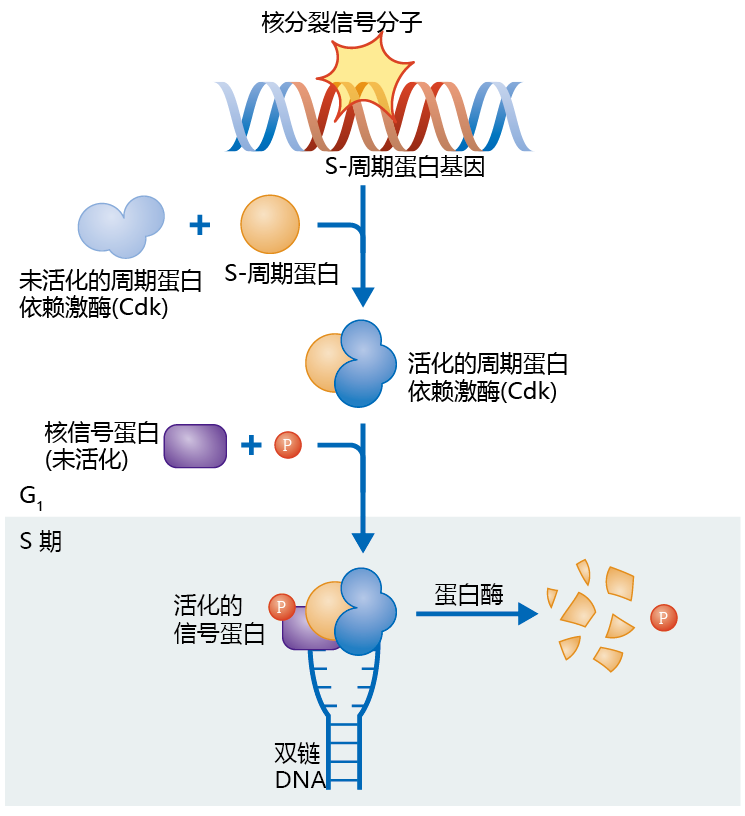
图4.4 细胞周期蛋白类触发DNA复制。胞外分裂信号附着到细胞,由此产生一个核分裂信号,从而诱导S-细胞周期的表达。S-细胞周期附着到周期蛋白依赖激酶(Cdk)激活这个酶并且引起其它蛋白质的磷酸化,从而促使细胞进入S期并启动DNA复制。一旦DNA复制开始,Cdk就被降解而失活。
一种叫做细胞周期蛋白的核蛋白质表达触发了DNA复制。胞外信号通过一些因子(例如生长激素)通过诱导核分裂信号蛋白的合成从而触发细胞周期。核分裂信号蛋白的种类随着胞外分裂信号的不同而不同。因此会有许多不同类型的S-周期蛋白会被表达。核分裂蛋白附着到一个启动子区域,DNA中的这个区域能够“打开”基因,从而诱导S-周期蛋白的表达(图4.4)。S-细胞周期附着到一个周期蛋白依赖激酶(Cdk)上面,这是一种论磷酸化酶,它一直存在于细胞核中。周期蛋白-Cdk复合体催化其它蛋白质的磷酸化,从而进入S期并启动DNA复制。蛋白质的磷酸化作为细胞信号的方式在真核生物中很常见。例如,在蠕虫中发现的信号通路,这是第一个发现的生殖关联到寿命的生化机制,下一章详细讨论。
一旦细胞进入S期,DNA的复制会刺激胞内蛋白酶的表达,蛋白酶分离周期蛋白和Ckd的复合体并将周期蛋白降解。最后一步保证在一个细胞周期中DNA只复制一次。
4.2.2 p53蛋白能将DNA复制阻止与G1和S期之间
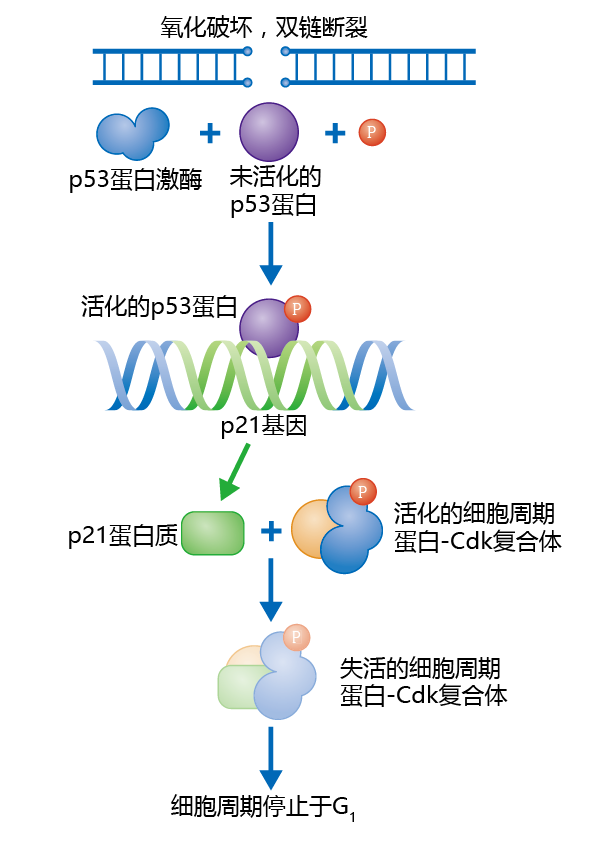
图4.5 G1期的p53检查点。当DNA的破坏被探测到时,例如超氧自由基的破坏(•O2−),修复机制活化p53蛋白激酶。磷酸化的p53蛋白附着到p21基因的调节区域,刺激p21基因的表达。然后p21蛋白结合到周期蛋白-Cdk复合体,使之失活并临时将细胞周期停止在G1期。一旦DNA被修复,蛋白酶降解p21蛋白,细胞进入S期。
为了保证DNA在合成过程中被准确复制,p53通路会阻止被破坏的DNA继续复制。例如由于受到自由基(本章后面会介绍自由基)的攻击而造成的某段DNA破坏。DNA修复机制探测到错误,开始修复DNA坏区,并且激活蛋白激酶,将核蛋白p53磷酸化。活化的p53附着到另一种蛋白质p21的启动子区域。p21通过负向阻止(negative inhibition)使周期蛋白-Cdk复合体失活并临时中止细胞周期(图4.5)。当修复完成后,蛋白酶被激活并分离p21和周期蛋白-Cdk复合体,细胞继续进行S期。p53通路在细胞衰老中起一个重要的角色,p53通路通过与端粒缩短的相互作用,在细胞衰老中发挥作用(看下面)。
4.2.3 DNA复制需要多种蛋白质参与
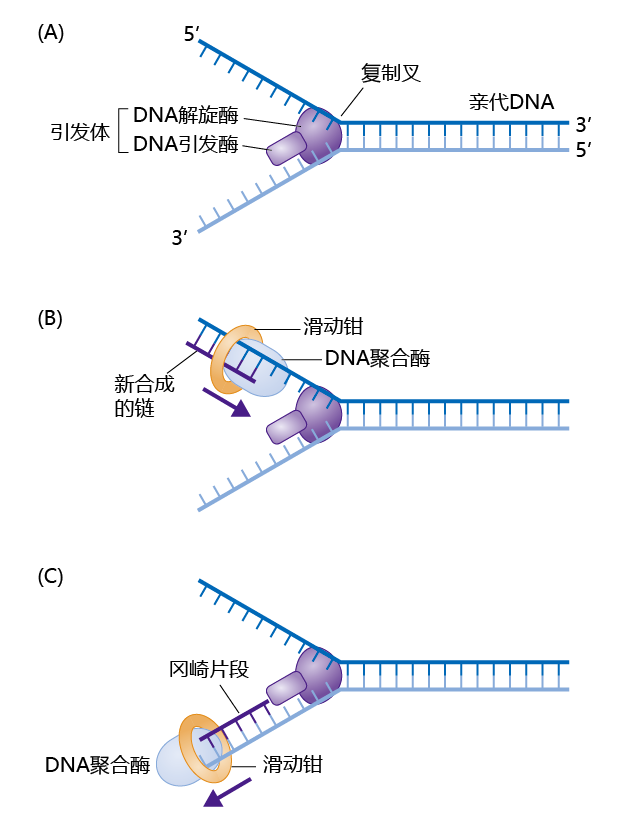
图4.6 DNA复制。(A)解旋酶将双链DNA分离成单链。DNA引发酶合成复制开始所需的RNA引物。 (B)在前导链模板上,DNA聚合酶在5′→3′方向上合成新链,此过程只需要一个引物即可。(C)在滞后链上,DNA聚合酶向相反方向移动,合成小的DNA片段,称作冈崎片段。冈崎片段被“倒缝”在母链上。滑动钳将DNA聚合酶卡在母链上,让它只能顺着模板链移动。
当DNA检查完成时,周期蛋白-Cdk复合体就被激活,DNA复制就可以开始。DNA复制需要好几种蛋白质按照一定的顺序来完成(图4.6A)。其中一种叫做解旋酶,需要ATP水解供能,将双链DNA解成单链,从而作为合成的模板。单链结合蛋白让这两条单链保持分离。另一种叫做引发酶(primase)的蛋白质合成一小段RNA分子用于引导复制。解旋酶和引发酶组成一个复制复合体,叫做引发体(primosome)。当DNA双链被分开并结合了RNA引物之后,DNA聚合酶就开始合成新链,它通过在新链的3'端逐个添加脱氧核糖核苷酸来进行。新链一直是从5′→3′方向合成,DNA聚合酶只会按照这个方向进行。
DNA复制从起始点向两边移动。因此,引发体分开的两条DNA单链模板分别是5′到3′方向和3′到5′方向(图4.6)。新合成的子链中,从5'到3'方向的那条连续的链叫做前导链,而从3'到5'方向的那条不连续的链(冈崎片段)叫做滞后链。由于DNA聚合酶将脱氧核糖核苷酸只添加到新链的3'端,因此前导链直接合成,不会间断。但是DNA聚合酶怎么合成滞后链呢?合成机制解决了这个问题,DNA聚合酶倒着移动,附着到滞后链模板上合成滞后链。当DNA聚合酶向后移动时,产生小的DNA片段,叫做冈崎片段(图4.6C)。此过程需要RNA引物,冈崎片段后面被DNA连接酶连接起来,形成一条完整的DNA链。
4.2.4 黏结蛋白和凝缩蛋白辅助控制染色体分离
在S期结束时,两条姐妹染色单体被黏结蛋白紧紧的绑定在一起(图4.7)。然而,染色质相对体积较大,而且杂乱,这样会妨碍它们高效的分离。因此在G2期装配出凝缩蛋白,在M期用以减少染色质的体积并提高分离过程。另一个关系到染色体正确分离的重要的复合体是着丝点。着丝点将两条姐妹染色体绑定在一起,并在此形成着丝粒,着丝粒将着丝点附着到纺锤体上。
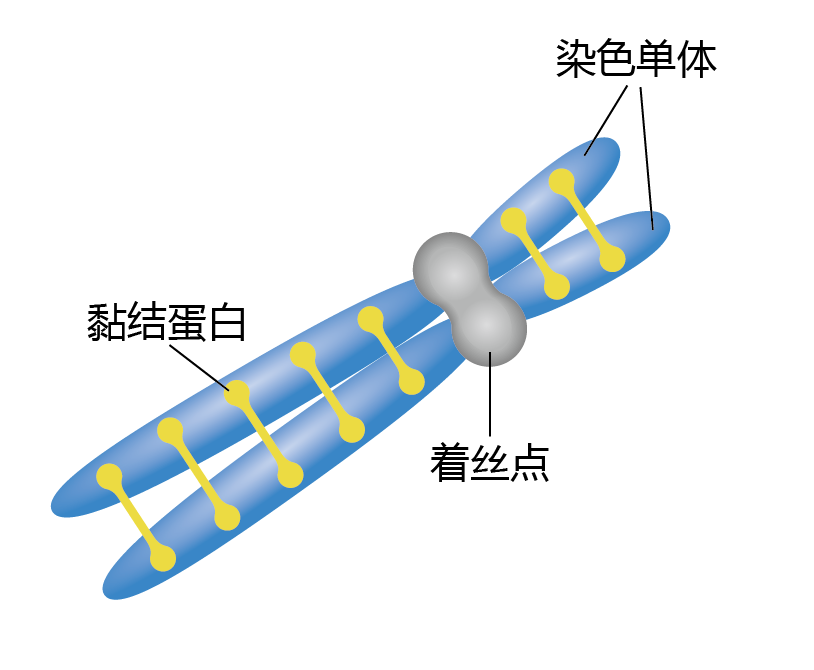
图4.7 在S期复制的姐妹染色单体。着丝点将两条姐妹染色单体绑定在一起。黏结蛋白也参与此过程。
4.2.5 中期到后期的过度是细胞周期最后的检查点
G2期到M期的过度是靠周期蛋白-Cdk复合体调节的。这个过程与图4.4描述的相似,但这里的周期蛋白是M-周期蛋白而不是S-细胞周期。一旦这个细胞周期控制点通过,M期就开始了,细胞周期中最后的调节步骤发生于中期到后期的过度。周期蛋白-Cdk复合体使用一个磷酸化和去磷酸化调控系统,因此中期到后期的过度会有一个蛋白水解过程。这个过程最初的蛋白质是后期促进复合物(APC),它是泛素连接酶家族成员。这个家族的很多酶都参与不正确折叠蛋白质的标记。
你在第九章将会看到,泛素连接酶在阿兹海默症和帕金森症的发育过程中起一个重要的作用。APC标记紧固蛋白的分解,紧固蛋白绑定到黏结蛋白,将姐妹染色单体绑定在一起。在中期之前,紧固蛋白阻止蛋白酶将姐妹染色单体分离。从黏结蛋白中去掉紧固蛋白就使得细胞中的蛋白酶可以分离姐妹染色单体了。然后纺锤丝开始收缩,染色单体分开。APC也标记残留周期蛋白,再次保证DNA只发生一次。从末期到胞质分裂细胞周期就不再受阻了,于是产生两个新的相同的子细胞。
4.2.6 Fully functional cells can exit the cycle at the G0 phase
多细胞生物只有接收到一个胞外信号才会启动细胞周期,这些物质叫做有丝分裂原(mitogen)。有丝分裂原成分各种各样,可以是激素,神经肽以及类固醇,它们可以距离被调控的细胞很远,也可以挨着。如果一个细胞没有在恰当的时间收到有丝分裂原信号,那么这个细胞就会终止细胞周期调控机制,进入一个G1期的修改版本,G0期。G0期的细胞不再分裂,但仍然具有原来的生理功能。当给予一个适当的有丝分裂原信号刺激时,有的成熟细胞可以重新返回到G1期。在人类的肠道细胞中发现,G0到G1的转换非常迅速而且很频繁(3到4天的转换速度),然而别的类型的细胞保持在G0期并且罕见分裂(肝细胞每年只分裂一次)。尽管一直处于分裂的细胞类型会超过不再分裂的细胞类型,但是多细胞生物永久停留在G0期的细胞数量更多。
表格4.1展示了不同G0期长度的细胞.全分裂的细胞在整个成熟阶段一直分裂;半分裂的细胞经常分裂;不分裂的细胞成熟之后不再分裂。G1到G0的转换机制并没有完全搞清楚,但在复制衰老(replicative senescence)中一定扮演一个重要的角色。
表格4.1 一直分裂,半分裂以及不再分裂的哺乳动物细胞
