
5.2 基因表达的调节
Expressing the correct gene, at the correct time, and in the correct amount is critical to the proper functioning of a cell. Disruption in the regulation of gene expression can lead to serious dysfunction within the cell and possible altered states of metabolism, which, in turn, can affect the organism‘s longevity. A problem facing biogerontologists in determining which of the 24,000 genes of the human genome affect longevity is that the type of gene expressed in a cell differs with the cell type. Hepatocytes (liver cells) and cardiac myocytes (heart cells) have radically different gene expression profiles for radically different functions, even though each cell contains the identical sequence of genomic DNA and uses the same transcription and translation processes to convert the genetic code of the DNA into 蛋白质. Nonetheless, as you learned in the previous chapter, because both cell types are somatic, both are mortal. These two facts—different gene expression in different cells and the universal mortality of somatic cells—have led many biogerontologists to suggest how unlikely it is that longevity reflects differences in a single gene, or even in groups of genes. Rather, many biogerontologists now believe that longevity reflects a complex interaction between gene expression and the processes that regulate gene expression, or lack of expression.
In this section, we examine the basic mechanisms that regulate gene expression. The processes that govern gene expression occur at virtually every step in conversion of the genetic code into a functioning 蛋白质. However, study of the genetic control of longevity is a very young science and has had time to elucidate only a few regulatory pathways in gene expression in sufficient detail to allow for discussion here. These studies have focused primarily on transcriptional regulation. Post-transcriptional control, the regulatory process occurring after RNA polymerase has bound to a gene‘s promoter, has been less well described in terms of its impact on the rate of aging and longevity. Therefore, we begin with a description of transcriptional regulation, then give a brief overview of post-transcriptional mechanisms—for which research has only just begun, or remains to be done—to describe their role in modulating longevity.
5.2.1 改变核小体结构可以控制基因表达
In eukaryotes, an efficient organization has evolved to deal with the enormous size of the DNA; if left in its linear form, DNA would easily occupy the entire volume of a cell (Figure 5.10). The DNA, RNA, and various 蛋白质 make up the material called chromatin. DNA is further organized into the chromosomes, bound to 蛋白质 called histones. Unfolded chromatin has the appearance of beads on a string. Each “bead” consists of DNA wound around a histone 蛋白质 called a histone octomer. Together, the wrapped DNA and histone 蛋白质 make up the nucleosome. The short segments or strings of DNA between nucleosomes are known as linker DNA.
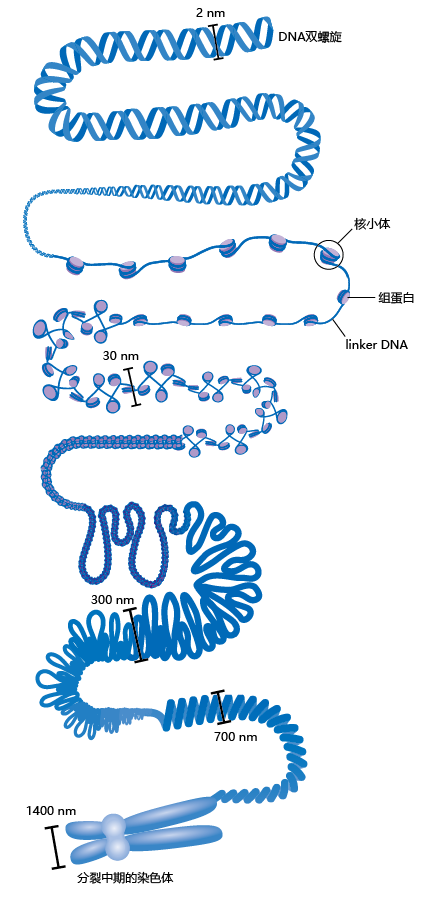
Figure 5.10 DNA packing in a eukaryotic chromosome.
Chromatin organization gives eukaryotes a significant 进化advantage over prokaryotes, because it protects the genes from inadvertent chemical reactions. However, the nucleosomes can also be a barrier to gene transcription, since they can block the TATA boxes (promoter regions) associated with genes. For transcription to go forward, the histones must be modified so that they loosen their grip on the DNA. This is accomplished by histone acetylation, a regulatory step that attaches acetyl groups (–COCH3) to certain amino acids in the histone. When histones are acetylated, they change shape so that they grip the DNA less tightly, particularly in the promoter region. As a result, transcription 蛋白质 can access the genes in the acetylated region (Figure 5.11). The opposite of histone acetylation, histone deacetylation, is discussed later in the chapter, as this mechanism appears to be important in the regulation of genes associated with longevity.
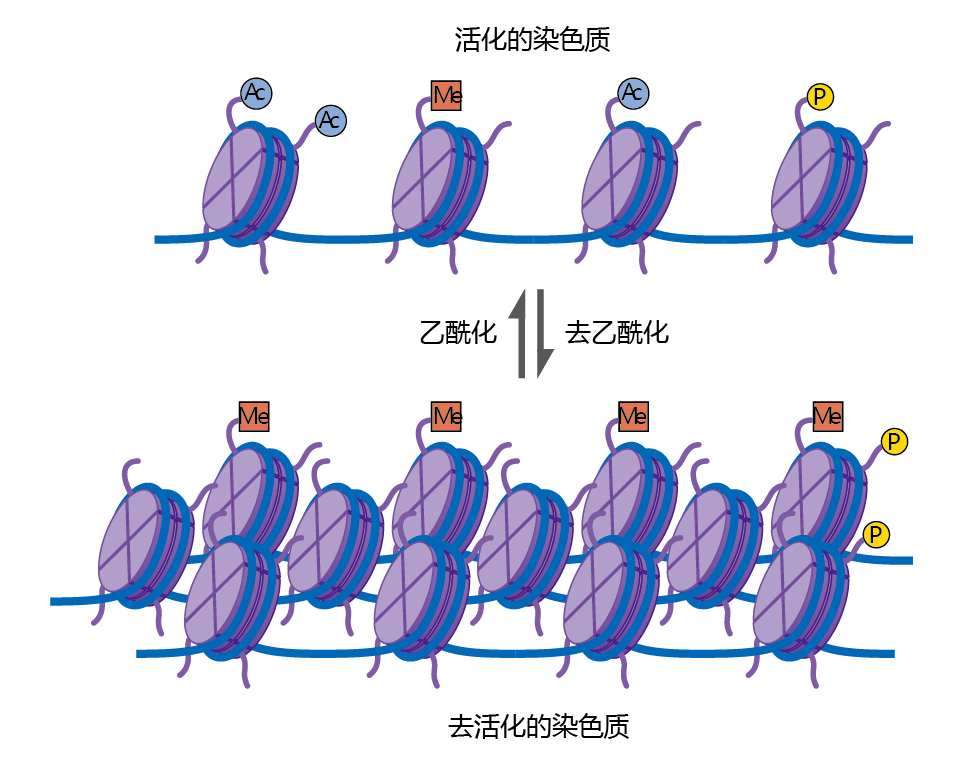
Figure 5.11 Histone acetylation and deacetylation. Before the initiation of gene transcription, the nucleosomes are tightly packed together. During histone acetylation, the addition of acetyl groups to the tails of the histones changes the charge on the DNA and “opens up” the pattern of nucleosome packing. The promoter regions now become more accessible to the binding 蛋白质 that initiate gene transcription. During deacetylation, the acetyl groups are removed from the histone tails, causing the nucleosomes to move closer together.
5.2.2 基因表达 is controlled by binding of 蛋白质 to DNA
One of the more important findings in genetics and cell biology was the discovery that the expression of genes can be turned on or off by the interaction of gene regulatory 蛋白质 with specific segments of noncoding DNA known as control elements (BOX 5.1). A eukaryotic 蛋白质-coding gene can contain two types of control elements: enhancers, which increase RNA transcription, and silencers, which decrease RNA transcription. A transcription factor that binds to an enhancer and stimulates transcription of a gene is called an activator. A transcription factor that binds to a silencer and inhibits transcription is called a repressor. Both activators and repressors may be hundreds or thousands of nucleotides away from the promoter. The regulation of transcription results mainly from binding of activators and repressors to enhancers and silencers and transmission of their effect to the 蛋白质 bound to the promoter regions. Control elements seem to have a significant role in determining the longevity of some species, as described later in this chapter.
| BOX 5.1 CONSERVATION OF GENETIC SWITCHES: FROM FRUIT FLY TO HUMAN |
| One of the greatest questions in biology after Watson and Crick‘s description of DNA structure was, “How can two cells of one animal contain identical DNA yet differ so radically in their functions?” For example, consider the differences between a human liver cell (hepatocyte) and muscle cell (myocyte). Among many other functions, hepatocytes detoxify harmful compounds in the blood by using a specialized set of enzymes. Myocytes also contain these enzymes, but the low level of expression of detoxifying 蛋白质 in these cells makes this pathway almost nonfunctional. On the other hand, actin and myosin, the major contractile 蛋白质 of the body, are expressed in muscle cells at levels millions of times greater than that observed in the liver—their function in the hepatocyte is limited to structures that hold the cell together. Prior to 1960, the differences in expression patterns among cells were thought to reflect a loss of genes during the differentiation process. This view was consistent with the observation that most metazoan cells lose the ability to divide after differentiation, although no empirical evidence describing a loss of specific genes existed. In the early 1960s, however, the pioneering work of François Jacob and Jacques Monod solved at least part of the mystery surrounding the regulation of gene expression. These Nobel laureates (along with André Lwoff) observed that, in the bacterium Escherichia coli, the enzyme β -galactosidase, which breaks the bond connecting glucose to galactose in the sugar lactose, was expressed only when lactose was present in the bacterium‘s growth medium. Using different mutants of E. coli, Jacob and Monod discovered that lactose influenced the binding of a 蛋白质 to DNA. When lactose was absent, the 蛋白质 was tightly bound to the DNA and prevented expression of β -galactosidase. Conversely, when lactose was present, the binding 蛋白质 fell off the DNA and β -galactosidase was expressed, leading to breakage of the bond between glucose and galactose. Jacob and Monod had discovered the first genetic switch, the lac repressor, a finding demonstrating that differences in gene expression among cells reflect a regulatory process rather than a difference in genes per se. Tens of thousands of genetic switches in virtually all species examined have been identified since Jacob and Monod's initial studies in bacteria. More importantly, subsequent research demonstrated that the DNA sequences in the binding sites of the genetic switches are highly conserved. For example, genes that determine the shape of the human body at the time of fetal development, known as the Hox genes, differ by less than one or two base pairs from the same genes found in the fruit fly. That is, the genes responsible for ensuring that our legs, arms, and fingers are in the proper position are not just similar but almost identical to the genes that determine the position of the wings, legs, and antennae of the fruit fly. Until this discovery, most scientists accepted the concept that differences between species reflected differences in the genes. That is, humans had very different genes than did the fruit fly. The discovery of the Hox genes changed this idea and suggested that, even though flies and humans took very different 进化 paths almost 500 million years ago, both species retained identical basepair sequences that regulate 蛋白质 expression. Differences between species do not reflect different genes; rather, they reflect differences in when and how much of a 蛋白质 is expressed. From a biochemical perspective, the fly and the human are identical. Indeed, the radical change in our understanding of 蛋白质 expression led Monod to state (and rightfully so), “What is true for E. coli is true for the elephant.” |
Activator 蛋白质 are expressed in response to hormones, metabolic products of hormones, amino acids, individual nutrients, and thousands of other types of molecules. When an activator 蛋白质 binds to the enhancer site, the promoter region of the gene bends to expose the TATA box for the binding of general transcription factors (Figure 5.12). DNA bending brings the bound activators closer to the promoter. The physical distortion in the DNA strand attracts a type of RNA polymerase called RNA polymerase II, as well as additional transcription factors and 蛋白质 that aid in transcription (well over a 100 蛋白质 are involved in this process). One such 蛋白质, known as a mediator, helps bind the enhancer and promoter regions. Together, RNA polymerase II, general transcription factors, and the mediator form the transcription initiation complex.
Repressor 蛋白质 can be as important as activator 蛋白质 to the regulation of gene transcription. Of the multiple mechanisms that eukaryotes use to repress gene transcription, four are most common: repressor 蛋白质 may (1) have a higher affinity for the enhancer site than does the activator 蛋白质; (2) mask the enhancer site by binding to a repressor site close to the activation site; (3) prevent formation of the transcription initiation complex; and (4) prevent formation of the gene activation complex altogether. This last is done indirectly through the process of histone acetylation-deacetylation. The first three mechanisms are shown in Figure 5.13. Later in this chapter, we discuss how the repression of gene transcription, also known as gene silencing, plays a significant role in the genetics of longevity.
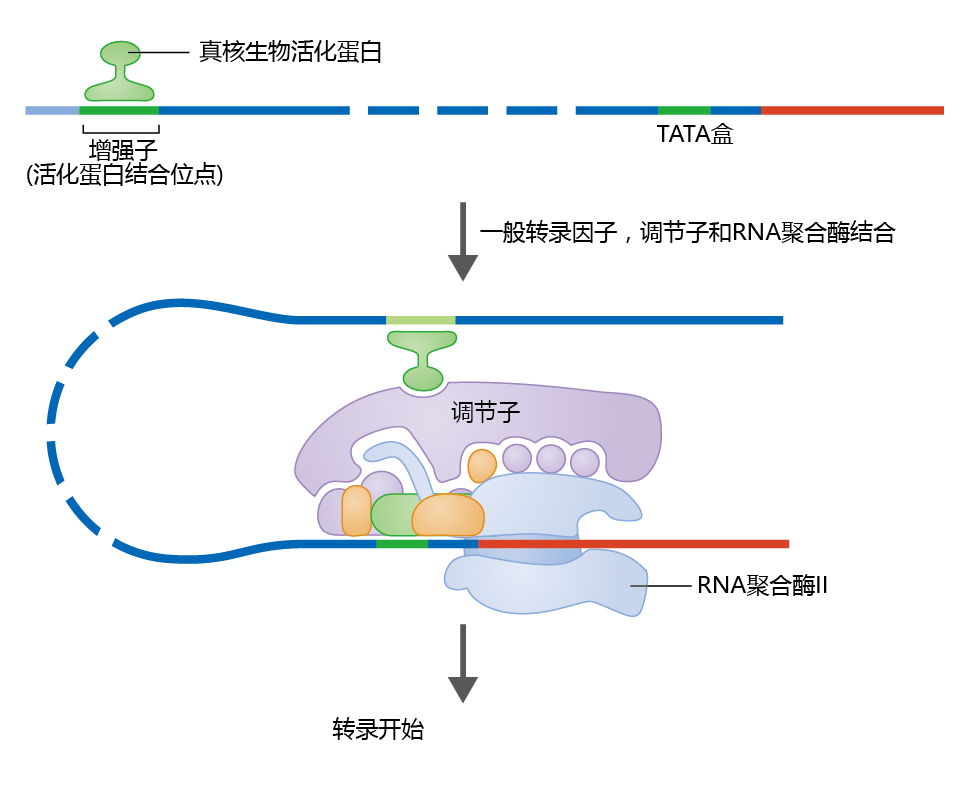 Figure 5.12 The action of activators and enhancers on gene transcription. Activator 蛋白质 bind to enhancer sequences in the DNA. The enhancer site, which may be thousands of nucleotides away from the promoter region, loops out and binds to the promoter region. Once the enhancer site comes within the proximity of the promoter, a mediator 蛋白质 helps the enhancer bind to the promoter region. The promoter region, bound enhancer site, and RNA polymerase form the transcription initiation complex.
Figure 5.12 The action of activators and enhancers on gene transcription. Activator 蛋白质 bind to enhancer sequences in the DNA. The enhancer site, which may be thousands of nucleotides away from the promoter region, loops out and binds to the promoter region. Once the enhancer site comes within the proximity of the promoter, a mediator 蛋白质 helps the enhancer bind to the promoter region. The promoter region, bound enhancer site, and RNA polymerase form the transcription initiation complex.
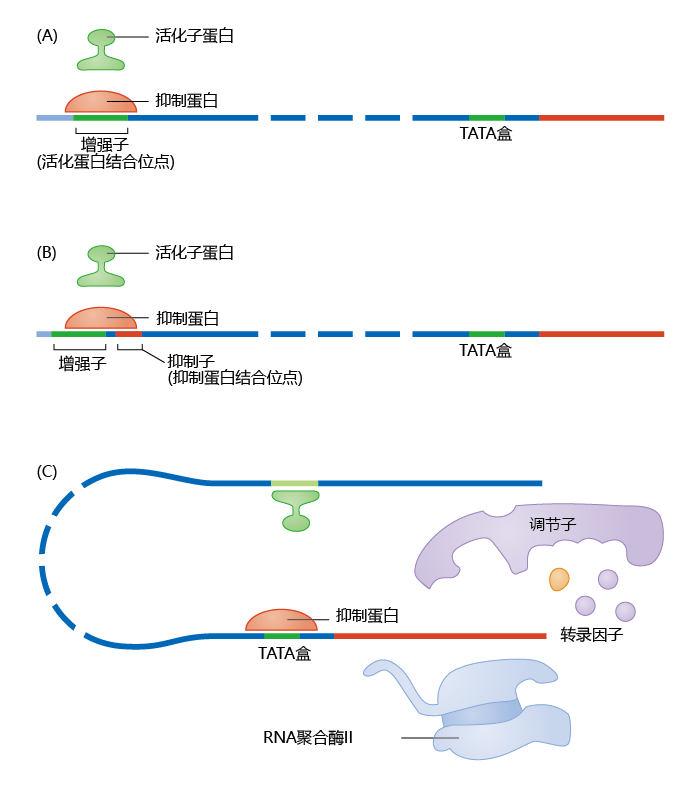
Figure 5.13 General mechanisms of gene transcription repression. (A) Competition between repressor and activator 蛋白质 for the same binding site. The repressor has a greater affinity for the binding site than does the activator. (B) Repressor 蛋白质 bind to a region near the enhancer site and prevent binding of the activator 蛋白质 by masking the enhancer site. (C) Repressor 蛋白质 prevent formation of the transcription initiation complex.
5.2.3 Post-transcriptional mechanisms can also control gene expression
While most gene regulation occurs at the transcriptional level, before RNA polymerase binds to the gene‘s promoter region, significant regulation occurs post-transcriptionally. Here we look briefly at two types of post-transcriptional control, alternative RNA splicing (Figure 5.14) and translational initiation, which will prove useful in evaluating the genetic control of the rate of aging and longevity.
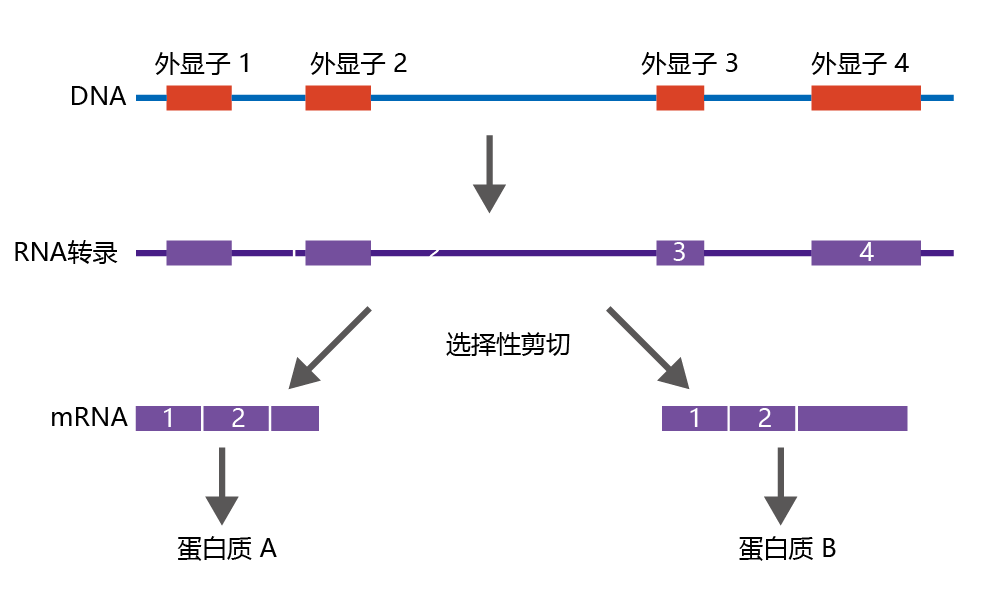
Figure 5.14 Alternative splicing can produce different mRNAs from the same gene.
Recall that introns are removed from the pre-mRNA transcript before mRNA becomes active, by the process of RNA splicing. The snRNAs that perform the splicing recognize the start and end of an intron by regions known as consensus sequences. However, these consensus sequences have significant plasticity and allow the snRNAs to begin splicing the intron at slightly different locations—the process of alternative RNA splicing. This results in changes in the nucleotide sequence of the finished mRNA, resulting in slightly different 蛋白质 with slightly different activity. That is, alternative RNA splicing allows the cell to make several variations of a common 蛋白质 from one gene, increasing the coding potential of the genome.
Regulation of alternative RNA splicing occurs by the binding of regulatory 蛋白质 to the pre-mRNA and can vary with cell type or developmental stage. For example, the Drosophila gene dsx encodes a 蛋白质 that is important to secondary sexual characteristics. If the RNA splicing results in six exons being translated, male sex characteristics are expressed. Conversely, if RNA splicing results in four exons in the mRNA, female characteristics are expressed. Thus, one gene results in two 蛋白质, suggesting different genotypes.
Cells can also control the amount of 蛋白质 being synthesized by regulating the initiation of translation. Before mRNA leaves the nucleus, RNA polymerase “caps” the 5′ end with a methyl group attached to a guanosine (G) nucleotide (see Figure 5.6). This capping is another way of distinguishing mRNA from other RNA fragments in the nucleus so that only mRNA moves through the nuclear pore complexes. The
methylguanosine cap also serves as a location of inhibition of translation. The cap is normally found very close to the first AUG (start) codon. Repressor 蛋白质 expressed in response to cellular signals recognize the methylguanosine cap and bind to the mRNA just before the AUG. Binding of the repressor 蛋白质 prevents the ribosome from translating the base sequence into the 氨基酸序列。 When cell conditions change, the repressor 蛋白质 is degraded and translation begins.